TOMATO
Lycopersicon esculentum Mill., family SolanaceaeThe tomato crop was produced on an estimated 395,500 acres in 1971 and valued at $444 million, making it second only to another vegetable of the same family, the potato (Solanum tuberosum L.), which was produced on 1,380,000 acres and valued at $626 million. Tomatoes are grown in almost every State, out of doors in season and in glass and translucent plastic houses for off-season markets. Recent innovations in mechanical harvesting and the breeding of cultivars that have improved shipping capabilities have caused considerable changes in the industry. Cold storage of fruit and its production under glass and translucent plastic, along with the ease of shipping tomatoes long distances now permit the public to have this vegetable on the food table throughout the year.
The plant is grown for its fruit, a fleshy berry, which is consumed fresh, canned, or used to produce juice, sauces, pastes, or powder. The seed yields 24 percent oil, which is used in salad oil, margarines, and soap (Purseglove 1968*). The number of seeds in a fruit may vary from 73 to 346 (Hafen and Stevenson 1956).
Plant:
The tomato, as it is grown in the United States, is a many-branched annual plant, 2 to 6 feet, at first erect but later becoming prostrate, with alternate manybranched leaves, 6 to 12 inches long, and clusters of 2 to 12 or more flowers. The plant is covered with short coarse hairs and has a glandular secretion with a characteristic unpleasant odor, particularly when bruised. It is cultivated in rows, 3 to 6 feet apart in the field, but usually tied up on strings when grown in greenhouses. Under commercial harvesting methods, cultivars are desired that set a maximum amount of fruit in a relatively short time, and the fruit retains its keeping qualities for several days both on the vine and after it is harvested.
Inflorescence:
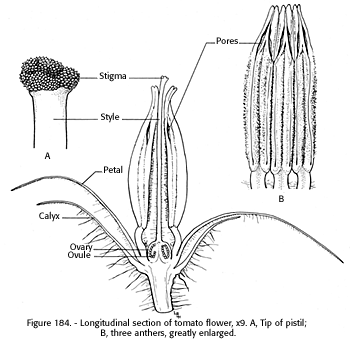
The inflorescence may arise terminally, opposite or between the leaves. The individual flower is about three-quarters of an inch in diameter with a 5- to 10-part green calyx, that clings to the fruit until it matures. There are usually six golden yellow petals that recurve as they expand. There are usually six stamens, which are united with their yellow anthers (fig. 184) to form a tube or cone about one-half inch long, that surrounds the pistil, and, with the recurved petals, gives the pendant flower a shooting star or rocket appearance. Depending upon cultivars and environmental conditions, the style may range from slightly shorter than the tip of the anthers to as much as 2 mm beyond the tip, terminating with a capitate, simple, narrow or somewhat bulbous stigma (Muller 1940). The flower is hermaphrodite, hypogynous, and regular.
The style elongates about the time the anthers begin to split at the terminal end and release their pollen into the styler tube. The stigma is receptive to its own or other pollen 1 or 2 days before anther dehiscence (Fink 1898), which favors cross-pollination. The stigma remains receptive for 4 to 8 days (Jones and Rosa 1928*). The construction of the anthers, delicately united with the filament, permits them to vibrate at the slightest touch and send a rain of pollen down the cone outlet and around the stigma.
Nectar secretion from tomato plants is apparently of little, if any, value in attracting bees. Schneck (1928) referred to "the absence of nectar" in the tomato flower. Neiswander (1954a, 1956) stated that the blossom "contains little or no nectar." Fink (1898) reported that bumble bees "gathered chiefly pollen'' from tomato flowers. Thus, if nectar is produced, a question that should be settled, it is of little significance in the relation of insect pollination of tomatoes. The pollen is more attractive to wild bees than honey bees.
[gfx] FIGURE 184. - Longitudinal section of tomato flower, x 9. A, Tip of pistil; B, three anthers, greatly enlarged.
Pollination Requirements:
Various tests in greenhouses have proven that the tomato flower is not self-pollinating. However, if the inflorescence is shaken, the pollen will fall from the anthers onto the stigma and fertilization will result. One pollen grain is needed for each seed, so many grains are needed on each stigma. Incomplete pollination results in misshapen fruit. Cool or cloudy weather retards pollen shedding (Stoner 1971). Growers of tomatoes in greenhouses use various types of vibrators or other devices each few days to shake the flower clusters (Beattie 1939; Bouquet 1919, 1924; Cottrell-Dormer 1945; Fletcher and Gregg 190 7; Hoffman 1958; Kerr and Kribs 1945; Lesley and Lesley 1939; Moore 1968; Neiswander 1954a, b, 1956; Ross 1963; Verkerk 1957; White 1918; Wittwer and Honma 1969). Moore (1968) obtained only 4.3 pounds of fruit from control plants in a plastic greenhouse; 6.6 pounds, from hormone treated plants; 8.8 pounds, from vibrator treated plants; and 9.8 pounds, from plants treated with both hormones and vibration. For maximum effectiveness, vibration must be repeated every 2 or 3 days.
Although Bailey and Lodeman (1895) concluded that bees in the greenhouse were of no value as pollinators of tomatoes, Neiswander (1954a, b, 1966) found that visits of honey bees increased fruit production even though the flowers had also received the shaking treatments. Marr and Hillyer ( 1968) showed that self-pollinated plants (in greenhouses) yielded less and had more misshapen fruit than crossed plants.
Jones (1916) observed 1.98 percent cross-pollination of tomatoes in New Jersey and estimated that an equal undetected amount of crossing occurred. This figure has been frequently used over the years, without regard to the insect pollinator population or variety under test. He stated that he "saw no insects," although others have associated cross- pollination with insects almost exclusively.
Tomato flowers in the open are usually considered to be sufficiently vibrated by wind currents to cause the pollen to fall onto the stigma and affect maximum set (Lesley and Lesley 1939). Wind is not a factor in transferring pollen from plant to plant (Currence and Jenkins 1942); however, if the weather remains calm or if the blossom is so situated on the plant that it is not vibrated by the wind its pollination would conceivably be prevented. Under such conditions, visitation by pollinating insects would be baneficial. Cross-pollination in the field is common although the percentage is usually low (Azzam 1960; Lesley 1924; Purseglove 1968*; Richardson and Alvarez 1957a; Rick 1947, 1949, 1950; Schneck 1928; Smith 1935; Soost and Rick 1957).
A factor favoring self-pollination of the tomato is the relatively long time that the stigma is receptive to pollen, from 1 to 2 days before anther dehiscence to 4 to 8 days after dehiscence (Smith 1935). Another factor is the length of the stigma. If the style is short and the stigma is surrounded by dehiscing anthers, eelfing after vibration is most likely. If the style is long or if it grows through the anther tube before pollen is shed, its likelihood of being cross-pollinated is increased. Jones and Rosa (1928*) stated: "In some varieties, however, and probably in some flowers of all varieties, the style elongates before the anthers dehisce, thus exposing the stigma to foreign pollen." Regardless of how the pollen is applied, the more pollen (within limits) the larger the fruit (Fink 1898), and the more symmetrical it is (Hoffman 1958).
Pollinators:
In greenhouses, the various types of mechanical vibrators are satisfactory pollinators. In the field, the wind vibrates the plants. Neither of these methods contributes to the pollination of male-sterile plants. Only insects can serve in this capacity (fig. 185). They also contribute to pollination of those plants or cultivars with styles that extend beyond the stigma. Currence (1944) showed that use of hybrids could increase yields by 20 percent, and he reported finding a male-sterile plant that set good crops from artificial pollination. Barrows and Lucas (1942) estimated the value of hybrid seed at $8 per ounce. This might be decreased if the grower could incorporate a seedling marker to aid in weeding out nonhybrids (Hafen and Stevenson 1956). Others (Hojby 1958, Kerr 1955, Oba et al. 1945, Roever 1948, and Wellington 1912) have shown the value of hybrid tomato production. Shifriss (1945) reported the production of hybrid tomato seeds, produced by the relatively inexpensive labor of college girls. Kerr (1955) associated greater numbers of seeds with larger and more rapid fruit development.
Where hand pollination is impractical, insects can be used. Richardson and Alvarez (1957a, b) considered the Halictid bee (Augochloropsis ignita Smith) the most effective pollinator in their area. Bullard and Stevenson (1953) considered neither houseflies, blowflies, nor honey bees of value under cheesecloth cages over six plants. Azzam (1960) observed few bumble bees on tomato flowers in Puerto Rico but several hundred Examalopsis glubosa F. bees.
It is generally known, however, that a few honey bees in such a cage do not act normally. Fletcher and Gregg (1907) hinted that honey bees might be used to distribute tomato pollen. Lesley and Lesley (1939) indicated that "bumble bees and other insects also assist." Neiswander (1954a, b, 1956) showed that honey bees can be of value as pollinators of tomatoes. Fink (1898) considered bumble bees to be effective pollinators. Rick (1950) suggested the use of "wild solitary bees" for cross- pollination of male-sterile tomatoes. Rick (1947) mentioned the value of insect pollinators of tomatoes and their protection from insecticides. Rick (1949) stated that at Riverside, Calif., Anthophora urbana Cresson was most common, but various species of solitary bees and a few species of bumble bees contributed to pollination of tomatoes. Schneck (1928) stated that bumble bees are fond of tomato flowers but that honey bees do not work them "probably because of the peculiar structure of the flower and the absence of nectar." The problem seems to be that wild pollinators in most areas are too scarce to have an impact on pollination of tomatoes from the production stand point.
Occasionally, honey bees visit tomato flowers, as was demonstrated in the greenhouse. Apparently, if they are sufficiently concentrated in a tomato-growing area, the competition could "force" them to visit tomato blossoms for pollen. Unless cultivars are found that produce nectar, there can be no insect pollination of male-sterile varieties for hybrid production.
If such cultivars are found, honey bees might be practical, or useful species of wild bees might be brought from Peru, the native homeland of the tomato, to provide adequate pollination. A new look should therefore be taken at current cultivars in which there has been incorporated new germ plasm to determine if nectar is being produced or if the flower has been changed in any other way that might affect pollinating insects.
Pollination Recommendations and Practices:
Because of current agricultural practices as well as the relative unattractiveness of tomato flowers to honey bees, many U.S. tomato fields are largely devoid of pollinating insects. Neiswander (1954a, 1956) con- cluded that honey bees should not replace vibrators in the greenhouse even though they increased production on an average of 1.1 pounds per plant on plants vibrated mechanically.
There are no recommendations for supplying pollinating insects to commercial fields, although the evidence indicates that if a heavy population of insect visitors could be established the effects would be beneficial.
LITERATURE CITED:
AZZAM, H.
1960. NATURAL CROSS-POLLINATION OF TOMATOES IN PUERTO RICO. Amer. Soc. Hort. Sci. Caribbean Region. Proc. 4: 85-86.BAILEY, L. H., and LODEMAN, E. G.
1895. INFORCING HOUSE MISCELLANIES. N.Y. (Cornell) Agr. Expt. Sta. Bul. 96, pp. 327-328.BARRONS, K. C., and LUCAS, H E.
1942. THE PRODUCTION OF FIRST GENERATION HYBRID TOMATO SEED FOR COMMERCIAL PLANTING. Amer. Soc. Hort. Sci. Proc. 40: 395-404.BEATTIE, J. H.
1939. GREENHOUSE TOMATOES. U.S. Dept. Agr. Farmers' Bul. 1431, 28 pp.BOUQUET, A. G. B.
1919. POLLINATION OF TOMATOES. Oreg. Agr. Expt. Sta. Bul. 158, 29 pp.____ 1924. ECONOMIC RESULTS IN THE POLLINATION OF GREENHOUSE TOMATOES. Oreg. Agr. Expt. Sta. Cir. 55, 16 pp.
BOLLARD, E. T., and STEVENSON, E. C.
1953. PRODUCTION OF HYBRID TOMATO SEED. Amer. Soc. Hort. Sci. Proc. 61: 451-458.COTTRELL-DORMER, W.
1945. AN ELECTRIC POLLINATOR FOR TOMATOES. Queensland Jour. Agr. Sci. 2: 157-169.CURRENCE, T. M.
1944. A COMBINATION OF SEMI-STERILITY WITH TWO SIMPLY INHERITED CHARACTERS THAT CAN BE USED TO REDUCE THE COST OF HYBRID TOMATO SEED. Amer. Soc. Hort. Sci. Proc. 44: 403-406.____ and JENKINS, J. M., JR.
1942. NATURAL CROSSING IN TOMATOES AS RELATED TO DISTANCE AND DIRECTION. Amer. Soc. Hort. Sci. Proc. 41 273-276.FINK, B.
1898. POLLINATION AND REPRODUCTION OF LYCOPERSICUM ESCULENTUM. Minn. Bot. Studies 1: 636-643.FLETCHER S. W., and GREGG, O. I.
1907 POLLINATION OF FORCED TOMATOES. Mich. Agr. Expt. Sta. Spec. Bul. 39: 2-10.HAFEN, L., and STEVENSON, E. C.
1956. NATURAL CROSS-POLLINATION IN TOMATO USING SEVERAL MALE-STERILE MUTANTS. Amer. Soc. Hort. Sci. Proc. 68: 433-436.HOFFMAN, I. C.
1958. POLLINATION PROBLEMS; HERE ARE WAYS TO OVERCOME POOR FRUIT-SET IN TOMATOES. Amer. Veg. Grower and Market Growers Jour. 6(3): 43-45.HOJBY, H. R.
1958. UTILIZATION OF HYBRID VIGOUR IN TOMATOES: POSSIBILITIES AND LIMITATIONS. So. African Jour. Agr. Sci. 1: 249-261.JONES, D. F.
1916. NATURAL CROSS-POLLINATION IN THE TOMATO. Science, n.s., 43: 509-510.KERR E. A.
1955. SOME FACTORS AFFECTING EARLINESS IN THE TOMATO. Canad. Jour. Agr. Sci. 35(3): 300-308.____ and KRIBS, W.
1945. ELECTRIC VIBRATOR AS AN AID IN GREENHOUSE TOMATO PRODUCTION. Queensland Jour. Agr. Sci. 2: 157-169.LESLEY, J. W.
1924. CROSS POLLINATION OF TOMATOES: VARIETAL DIFFERENCES IN AMOUNT OF NATURAL CROSS POLLINATION IMPORTANT FACTOR IN SELECTION. Jour. Hered. 15: 233-235.____ and LESLEY, M.
1939. UNFRUITFULNESS IN THE TOMATO CAUSED BY MALE STERILITY. Jour. Agr. Res. 58: 621-630.MARR, C., and HILLYER, I. G.
1968. EFFECT OF LIGHT INTENSITY ON POLLINATION AND FERTILIZATION OF FIELD AND GREENHOUSE TOMATOES. Amer. Soc. Hort. Sci. Proc. 92: 526-530.MOORE, E. L.
1968. OBTAINING FRUIT SET OF PLASTIC GREENHOUSE TOMATOES. Miss. Agr. Expt. Sta. Bul. 768, 8 pp.MULLER, C. H.
1940. A REVISION OF THE GENUS LYCOPERSICUM. U.S. Dept. Agr. Misc. Pub. 382, 28 pp.NEISWANDER, R. B.
1954a. HONEY BEES AS POLLINATORS OF GREENHOUSE TOMATOES. Gleanings Bee Cult. 82: 610-613.______ 1954b. HONEY BEES AS TOMATO POLLINATORS. Ohio Veg. and Potato Growers' Assoc. 39th Ann. Proc., pp. 96, 98,100,102, 104, 106, 108.
______ 1956. POLLINATION OF GREENHOUSE TOMATOES BY HONEY BEES. Jour. Econ. Ent. 49: 436-437.
OBA, G. I., RINER, M. E., and SCOTT, D. H.
1945. EXPERIMENTAL PRODUCTION OF HYBRID TOMATO SEED. Amer. Soc. Hort. Sci. Proc. 46: 269-276.RICHARDSON, R. W., and ALVAREZ, L. E.
1957a. POLLINATION RELATIONSHIPS AMONG VEGETABLE CROPS IN MEXICO. 1. NATURAL CROSS-POLLINATION IN CULTIVATED TOMATOES. Amer. Soc. Hort. Sci. Proc. 69: 366-371.______ 1957b. [NATURAL CROSSING IN THE TOMATO.] Agr. Tecnologia Mex. 3: 18, 44-45. [In Spanish, English summary.]
RICK, C. M.
1947. THE EFFECT OF PLANTING DESIGN UPON THE AMOUNT OF SEED PRODUCED BY MALE STERILE TOMATO PLANTS AS A RESULT OF NATURAL CROSS-POLLINATION. Amer. Soc. Hort. Sci. Proc. 50: 273-284.______ 1949. RATES OF NATURAL CROSS-POLLINATION OF TOMATOES IN VARIOUS LOCALITIES IN CALIFORNIA AS MEASURED BY THE FRUITS AND SEEDS SET ON MALE-STERILE PLANTS. Amer. Soc. Hort. Sci. Proc. 54: 237-252.
______ 1950. MALE STERILE TOMATOES. Calif. Agr. 4(4): 7, 12.
ROEVER, W. E.
1948. A PROMISING TYPE OF MALE-STERILITY FOR USE IN HYBRID TOMATO SEED PRODUCTION. Science 107: 506.ROSS, R. C.
1963. TRUSS VIBRATION INCREASES TOMATO PROFITS. Agr. North Ireland 37(12): 378.SCHNECK, H. W.
1928. POLLINATION OF GREENHOUSE TOMATOES. N.Y. (Cornell) Agr. Expt. Sta. Bull 470, 60 pp.SHIFRISS, O.
1945. HYBRID TOMATO. South. Seedsman 8(4): 15.SMITH, O.
1935. POLLINATION AND LIFE HISTORY STUDIES OF THE TOMATO LYCOPERSICON ESCULENTUM MILL. N.Y. (Cornell) Agr. Expt. Sta. Mem. 184, 16 pp.SOOST, R. K., and RICK, C. M.
1957. EFFECT OF VARIETIES OF POLLEN AND OVULE PARENTS ON NATURAL CROSS-POLLINATION OF TOMATOES. Amer. Soc. Hort. Sci. Proc. 70: 357-365.STONER, A. K.
1971. COMMERCIAL PRODUCTION OF GREENHOUSE TOMATOES. U.S. Dept. Agr., Agr. Handb. 382, 32 pp.VERKERK, K.
1957. THE POLLINATION OF TOMATOES. Netherlands Jour. Agr. Sci. 5(1): 37-54.WELLINGTON, R. [A.]
1912. INFLUENCE OF CROSSING IN INCREASING THE YIELD OF THE TOMATO. N.Y. (Geneva) Agr. Expt. Sta. Bul. 346: 423 - 442.WHITE, T. H.
1918. EXPERIMENTS WITH FERTILIZERS ON GREENHOUSE CROPS. Md. Agr. Expt. Sta. Bul. 222, pp. 77-101.WITTWER, S. H., and HONMA, S.
1969. GREENHOUSE TOMATOES - GUIDELINES FOR SUCCESSFUL PRODUCTION. Michigan State University Press, East Lansing, 95 pp.