STRAWBERRY
Fragaria X ananassa Duch, family RosaceaeThe estimated acreage of U S. strawberries in 1971 was 51,000, and the crop was valued at $116 million.
Oregon was the leading grower of strawberries, with 11,000 acres, followed by California (8,300 acres), Michigan (5,600), Washington (4,100 acres), and North Carolina (1,900 acres). Numerous other States produced more than 1,000 acres. California led with 151,500 tons, followed by Oregon (41,650 tons), Washington (13,350 tons), Michigan (12,300 tons), and Florida (7,600 tons). The numerous cultivars have changed rapidly over the last few years (Scott 1971). Much of the material herein is drawn from the excellent reference by Darrow (1966).
Plant:
The strawberry plant is a stemless, low creeping, and usually perennial herb that may live for many years, although it is sometimes grown as an annual in the South (Shoemaker 1955). Some cultivars are evergreen and others tend to be deciduous, depending upon the area in which they are grown. The trifoliate leaves form a blanket cover of the ground from a few inches to 2 feet deep, which shelters the fruit. The creeping runners occasionally produce roots and inflorescences at the leaf bases.
The ripe fruit is 1 to 2 inches long and light red to dark red when ripe. It is an ovoid aggregate of achenes or one-seeded fruitless around a receptacle that accumulates sugars and vitamins and ripens like a true, fleshy fruit. Each achene contains a single ovule and can therefore be considered an individual unit. Yet, if the stigma of the achene is not pollinated or if it is removed soon after pollination, there is no growth on the receptacle. The weight of the strawberry is roughly proportional to the number of fertilized ovules (Nitsch 195O).
Reproduction is almost exclusively with rooted runners, even though the seeds are viable.
Inflorescence:
The strawberry flower cluster is a series of double-branching parts bearing a flower in the crotch of each branch. The flower in the first crotch is termed the primary flower, the two in the next two crotches are termed secondary flowers, the next four are tertiary flowers, the next eight are the quarternary, and the next 16, if they develop, are the quinary flowers. The primary flower opens first and usually produces the largest berry (fig. 175) (Shoemaker 1955).
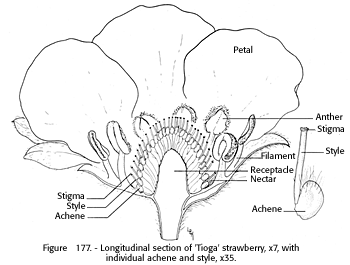
The individual flower is whitish (fig. 176), 1 to 1l/2 inches across, and usually composed of about 5 to 10 green sepals, five oval petals, numerous styles, and two to three dozen stamens arranged in three whorls. When the stamens contain viable pollen, they are a deep gold. Nectar is secreted by the receptacle and held at the base of the stamens next to the outer row of pistils.
The flowers of all current commercial cultivars are hermaphrodite. Clones that are only staminate or only pistillate may appear in the wild or in some seedling populations (fig. 177). The hermaphrodite flowers set fruit somewhat in proportion to the extent of pistillateness, that is, the higher the percentage of pistillate flowers, the more fruit the plant produces.
The stigmas are receptive before pollen of the same flower is available, which encourages cross-pollination. Sometimes, flowers that have pollen-laden anthers appear to set fruit far better when cross- pollinated than when fertilized with their own pollen. The pollen is mature before the anthers dehisce, but dehiscence does not occur until after the flower opens and the anthers dry a short while. This causes them to dehisce under tension so that the pollen is thrown onto some of the pistils. It can remain viable for several days, but some flowers are dried and shrunken on the second day after opening (Connor 1970); therefore, it is no longer of value to the flower. No complete self-incompatibility exists in present-day cultivars.
The fruit of the first blossom to open is referred to as the primary berry and is usually the largest. The second flower to open is the secondary flower, and the fruit it produces is usually second in size. Fruits from later flowers are usually smallest. Darrow (1966) stated that Valleau (1918) found 382 seeds in primary berries, and 224, 151, and 92 seeds in the succeeding berries. Gardner (1923) recorded 518 pistils on one primary flower but only 83 pistils for the last flowers of the plant under his study. There can be less but never more achenes (fruitless) than there are pistils.
[gfx] FIGURE 175. - Cluster of 'Midway' strawberries in different stages of developement.
FIGURE 176. - Strawberry blossoms, buds, and leaves.
FIGURE 177. - Longitudinal section of 'Tioga' strawberry, x 7, with individual achene and style, x 35.
Pollination Requirements:
Flowers without stamens were common in earlier cultivars, and no fruit setting resulted unless pollen was brought from staminate flowers (Darrow 1927, 1937). Continued breeding and selecting has resulted in the hermaphrodite flowers in all commercial cultivars. However, hermaphrodite flowers may not be completely self-fertilizing. The stamens are so placed that when they crack open they readily scatter pollen onto many, but not necessarily all, of the pistils. Pollination of all of the pistils of a flower is necessary for maximum berry size. If all pistils are fertilized, a perfectly shaped berry should develop. If few are fertilized, an irregularly shaped berry or "nubbin," sometimes only one- fifth the size of well-fertilized berries, will develop.
Allen and Gaede (1963) studied fruit-setting of 'Shasta' strawberries in the greenhouse and showed that plants caged and undisturbed by man, insects, or breezes set no fruit; those uncaged and undisturbed set 20 percent; those uncaged but receiving wind from a fan over them set 77 percent; whereas those that were caged, but brush pollinated daily, set 97 percent of the flowers. This finding indicated that the plants alone set few fruits, and wind has some effect, but insects may be more important than wind as pollinating agents. Couston (1966) also noted that malformation of berries was greater when adverse weather occurred at flowering time. He also obtained more number one berries from exposed plants than from plants caged to prevent insect visitation, indicating that insect pollination increased production.
Free (1968a) compared production of plots caged to exclude pollinating insects, plots caged with a colony of honey bees in each cage, and open plots. The cages without bees yielded the lowest percentage set, 55 percent as against 65.5 percent for the cage with bees. They also yielded the smallest berries, 6.7 g per berry in the cage without bees as against 8.3 and 8.4 g in the cages with bees and open plots, and the highest percentage of malformed berries, 48.6 percent in the no bee cages, 20.7 percent in the bee cage, and 15.4 percent in the open plots. Howitt et al. (1965) also associated strawberry fruit deformity with faulty pollination. Hughes (1961) noted that excluding pollinating insects resulted in decreased yield and malformed fruit. Kronenberg (1959) and Kronenberg et al. (1959) listed insufficient pollination by bees as one cause of poor fruit set.
Darrow (1966) stated that when the first flowers of perfect- flowered cultivars open and set well, but later flowers only partially set or do not set at all, natural sterility is the primary cause. However, he said that if the first flowers develop into nubbins, and yet later flowers produce good berries, the poor development is probably due to inadequate pollination.
Connor and Martin (1973) made the interesting observation that stamen height ranged from 2.5 mm in 'Surecrop' to 5.2 mm in 'Early Midway', and the flowers with the shorter stamens benefit most from insect pollination. Based upon their studies of 11 cultivars, they reported the following: "Self-pollination is responsible for development of 53 percent of the achenes; wind motion increased this development to 67 percent and insect pollination increased it to 91 percent."
Pollinators:
Many types of insects visit strawberry flowers, including flies, beetles, thrips, butterflies, and various bees; however, only the bees are of real consequence in transferring pollen effectively without injuring the flower parts. If wild bees are not plentiful so that the flower obtains the 16 to 25 bee visits recommended by Skrebtsova (1957), honey bees can be provided. Honey bees show preference for some cultivars over others, and they are not too strongly attracted to strawberries. However, this can be overcome with saturation pollination, or overstocking the area with colonies so the competing nectar and pollen are removed (fig. 178).
Although the strawberry blossom produces nectar and usually pollen, it is not overly attractive to honey bees. Also, different cultivars are visited by bees to different degrees, but none have been reported to be highly attractive. Free (1968a) stated that honey bee visits tend to be limited to good weather. Allen (1937) stated that when bees visited the strawberry blossoms, fruit production was increased, but the blossoms were not as popular with bees as one might wish.
In studying the activity of honey bees on strawberries, Free (1968b) found that although bees sometimes landed on the petals of a flower and approached the nectary from the side, they nearly always proceeded to walk over the stigmas. Some bees collected mostly nectar but also some pollen; however, some bees collected pollen deliberately. He stated, "Such bees either walked round the ring of anthers and scrabbled for pollen while doing so, or stood on the central stigmas and pivoted their heads and parts of their thoraxes over the ring of anthers. Some bees scrabbling for pollen also collected nectar." In either case, honey bees in sufficient numbers should be effective pollinators.
Skrebtsova (1957) studied visitation of strawberry flowers by honey bees. She noted that the bees showed preference for some cultivars over others, but concluded that each flower should receive at least 16 to 20 visits. More visits resulted in heavier berries; 16 to 20 visits resulted in berries weighing 5.36 g, and 21 to 25 visits produced berries that averaged 8.13 g. Flowers pollinated at the most receptive time, the time of fullest development of the reproduction organs, produced berries 13.3 to 58.3 percent heavier than those pollinated before or after this time. Later, Skrebtsova (1958) recommended saturated pollination by bees to produce the maximum crop of highest quality berries.
Moore (1964) noted that strawberry flowers are receptive up to 7 days after opening. Darrow (1966) said 10 days in cool weather, but the number of seeds per berry was reduced in late-pollinated flowers. As previously stated, Connor (1970) reported that many flowers were dried and shrunken on the second day after opening. The best time for pollination seems to be during the first 1 to 4 days after the flower is open. Darrow (1966) noted that reaction to pollination is rapid, within 24 to 48 hours the petals fall and the pistils dry up. Connor (1970) did not distinguish between pollinated and other flowers.
Fletcher (1917) stated that 90 percent of the pollination of strawberries was performed by insects and that honey bees accounted for 90 percent of this activity. Lounsberry (1930) stated that when bee forage was marginal, the bees worked strawberry blossoms feverishly. In Russia, Shashkina (1950) concluded that wind was not a good pollen vector but that flies were the principal vectors in the Moscow area.
Mommers (1961) showed that honey bees increased production in the greenhouse. This was supported by Bonfante (1970). Muttoo (1952) stated that location of an apiary near a strawberry plot increased the average production of berries from 840 to 1,225 pounds per acre. Petkov (1963) stated that only 31 to 39 percent of flowers isolated from bees developed fruit compared to 55 to 60 percent of those freely visited by pollinating insects. Furthermore, the isolated flowers developed 60 to 65 percent culls compared to 14 to 17 percent culls from bee-visited flowers. The average weight of fruit from the isolated flowers was only a third of that from the bee-visited flowers.
Petkov (1965) stated that over a 4-year period of observation bees accounted for 50 to 78 percent of the flower visitors (in Bulgaria). Pammel (1930, p. 922) noted that the flowers at Ames, Iowa, were visited by honey bees and by species of Halictus.
Moore (1969) studied the effects of caging and bees on strawberries over a 4-year period. He concluded that caging, which reduced yields 41, 32, 59, and 71 percent, respectively, for the years 1965-68, was " . . . due to incomplete pollination as a result of excluding insects." While pollinating insects were shown to be unessential for fruit set, the maximum yields and fruit size are only realized under conditions of adequate and active insect pollinators. Anderson (1969) cited numerous references relating to strawberry pollination.
[gfx] FIGURE 178. - Honey bee collecting pollen from strawberry blossom. In the process thorough pollination of the flower is assured.
Pollination Recommendations and Practices:
Free (1968a) discouraged the rental of bees " . . . unless the plantation is large." Darrow (1966) did not consider supplemental pollination, although he showed the need for insect visitation. Mommers (1961) recommended the use of bees on strawberries in greenhouses. Moore (1969) stated that "growers may someday have to provide colonies of honey bees for their plantations." Jaycox (1970) recommended one strong colony per 2 acres, with the bees in two or more groups on opposite sides of 10- to 50-acre fields. Unfortunately, as Connor (1970) pointed out, no specialist can wisely point to a strawberry field and state that the field is either well pollinated or needs additional pollination activity.
In general, strawberry growers do not take steps to provide additional pollination, nor even give much consideration to the local pollinator population in the field, although the evidence shows that visitation by pollinating insects is highly beneficial. The acreage devoted to strawberries on most farms is small compared to the foraging range of bees from an apiary, although some fields of 200 acres are in this crop. Thus, many acres of plants around a strawberry field can be more attractive to honey bees than the strawberry flowers. To overcome this competition would call for saturation pollination, or the placement of many colonies for each acre of strawberries, possibly 5 to 10 or even more. For a commercial grower who desires the maximum in perfect berries as well as in volume, saturation pollination should be profitable.
LITERATURE CITED:
ALLEN, M. Y. 937.
1937. EUROPEAN BEE PLANTS. 148 pp. The Bee Kingdom League, Alexandria, Egypt.ALLEN, W. W., and GAEDE, S. E.
1963. STRAWBERRY POLLINATION. Jour. Econ. Ent. 56: 823-825.ANDERSON, W.
1969. THE STRAWBERRY, A WORLD BIBLIOGRAPHY. 731 pp. Scarecrow Press Inc., Metuchen, N.J.BONFANTE, S.
1970. [POLLINATION OF THE STRAWBERRY.] Conv. Naz. della Fragola, Atti. Cesena (Italy) 4: 291-296. [In Italian.]CONNOR, L. J.
1970. STUDIES OF STRAWBERRY POLLINATION IN MICHIGAN. In The Indispensable Pollinators, Ark. Agr. Ext. Serv. Misc. Pub. 127, pp. 157-162.______and MARTIN, E. C.
1973. COMPONENTS OF POLLINATION OF COMMERCIAL STRAWBERRIES IN MICHIGAN. HortScience 8: 304-306.COUSTON, R.
1937. STRAWBERRY IMPROVEMENT U.S. Dept. Agr. Yearbook 1937: 445-495.DARROW, G. M.
1927. STERILITY AND FERTILITY IN THE STRAWBERRY. Jour. Agr. Res. 34: 393-411DARROW, G. M.
1937. STRAWBERRY IMPROVEMENT. U.S. Dept. Agr. Yearbook1937: 445-495______ 1966. THE STRAWBERRY. 447 pp. Holt, Rinehart and Winston, New York, Chicago, and San Francisco.
FLETCHER, S. W.
1917. THE STRAWBERRY IN NORTH AMERICA. HISTORY, ORIGIN, BOTANY AND BREEDING. 325 pp.The Macmillan Co., New York.FREE J. B.
1968a. THE POLLINATION OF STRAWBERRIES BY HONEYBEES. Jour. Hort. Sci. 43: 107-111._______ 1968b. THE FORAGING BEHAVIOUR OF HONEYBEES (APIS MELLIFERA) AND BUMBLEBEES (BOMBUS spp.) ON BLACKCURRANT (RIBES NIGRUM), RASPBERRY (RUBUS IDAEUS) AND STRAWBERRY FRAGARIA x ANANASSA) FLOWERS. Jour. Appl. Ecol 5: 157-168.
GARDNER, V. R.
1923. STUDIES IN THE NUTRITION OF THE STRAWBERRY. Mo. Agr. Expt. Sta. Res. Bul. 57, 31 pp.HOWITT, A. J., PSHEA, A., and CARPENTER, W. S.
1965. CAUSES OF DEFORMITY IN STRAWBERRIES EVALUATED IN A PLANT BUG CONTROL STUDY. Mich Agr. Expt. Sta. Quart. Bul. 48(2): 161-166.HUGHES, H. M.
1961. PRELIMINARY STUDIES ON THE INSECT POLLINATION OF SOFT FRUITS. Expt. Hort. 6: 44.JAYCOX, E. R.
1970. POLLINATION OF STRAWBERRIES. Amer. Bee Jour. 110: 176-177.KRONENBERG, H. G.
1959. POOR FRUIT SETTING IN STRAWBERRIES. I. CAUSES OF A POOR FRUIT SET IN STRAWBERRIES IN GENERAL. Euphytica 8: 47-57.______BRAACK, J. P., and ZEILINGA, A. E.
1959. POOR FRUIT SETTING IN STRAWBERRIES. II. MAL-FORMED FRUITS IN JUCUNDA. Euphytica 8: 245-251.LOUNSBERRY, C. C.
1930. VISITS OF HONEY BEES. In Pammel, L. H., and King, C. M., Honey Plants of lowa, lowa Geol. Survey Bul. 7 pp. 1106-1109.MOMMERS, J.
1961. [POLLINATION OF STRAWBERRIES UNDER GLASS.] Bijenteelt 63: 138-139. [In Dutch]MOORE, J. N.
1964. DURATION OF RECEPTIVITY TO POLLINATION OF FLOWERS OF THE HIGH-BUSH BLUEBERRY AND THE CULTIVATED STRAWBERRY. Amer. Soc. Hort. Sci. Proc. 85: 295-301.______ 1969. INSECT POLLINATION OF STRAWBERRIES. Amer. Soc. Hort. Sci. Proc. 94: 362-364.
MUTTOO R. N
1952. BEES PRODUCE MORE FRUIT. Indian Bee Jour. 1(7): 118-119.NITSCH, J. P.
1950. GROWTH AND MORPHOGENESIS OF THE STRAWBERRY AS RELATED TO AUXIN. Amer. Jour. Bot. 37: 211-215.PAMMEL, L. H.
1930. BRIEF HISTORY OF INVESTIGATIONS OF FLOWER POLLINATION. In Pammel, L. H., and King, C. M., Honey Plants of Iowa, Iowa Geol. Survey Bul. 7, pp. 850-966.PETKOV, V. G.
1963. THE SHARE OF BEES IN THE POLLINATION OF STRAWBERRY PLANTS. In 19th Internatl. Apic. Cong. Proc., Prague, pp. 94-95.______ 1965. [STUDIES ON THE PARTICIPATION OF BEES IN STRAWBERRY POLLINATION.] Gradinar. i Lozar. Nauka 2(4): 421-431. [In Bulgarian, German and Russian summaries.] (Abstract in Plant Breed. Abs. 36: 4981, P. 625. 1966.)
SCOTT. D. H.
1971. STRAWBERRIES - CHANGING PRODUCTION PATTERNS. Amer. Fruit Grower 91(1): 13-15, (2): 18-19.SHASHKINA, L. M.
1950. [CROSS-POLLINATION OF STRAWBERRIES (FRAGARIA GRANDIFLORA EHRH.).] Agrobiologiya 5: 45-47. [In Russian.]SHOEMAKER, J. S.
1955. SMALL FRUIT CULTURE. 441 PP. McGraw-Hill Publishing Co., Inc., New York.SKREBTSOVA, N. D.
1957. [ROLE OF BEES IN POLLINATION OF STRAWBERRIES.] Pchelovodstvo 34(7): 34-36. [In Russian.]______ 1958. [EFFECTIVENESS OF THE SATURATED POLLINATION OF STRAWBERRIES BY BEES.] Biull. Nauchno-Tekh. Inform (Nauchno-Issled. Inst. Pchelovod. ) (3-4): 25 - 26. [ In Russian. ]
VALLEAU, W. D.
1918. HOW THE STRAWBERRY SETS ITS FRUIT. Minn. Hort. 46: 449-454.