PEANUT
Arachis hypogaea L., family LeguminosaePeanuts are also known as goobers, groundnuts, and pincers. Approximately 1.5 million acres were planted to peanuts in 1969, and the value of the crop was $311.3 million. This frost-sensitive plant is grown in the southeastern and southern States, primarily for its seed, the peanut, which is a pea and not a true nut. The foliage is sometimes used for livestock feed.
Plant:
The peanut plant is an erect to spreading branched annual, 10 to 20 inches tall, that is cultivated in rows about 1 1/2 to 3 feet apart. The seeds are planted in the spring after all danger of frost is past, and the crop is usually harvested before frost in early fall. The plant has a primary taproot with weak laterals that permit easy removal of the entire plant from the soil. The leaves have four leaflets, 1 1/2 to 2 1/2 inches long. The seeds develop just below the surface of the soil, but they are attached to the branches near the base of the plant.
Inflorescence:
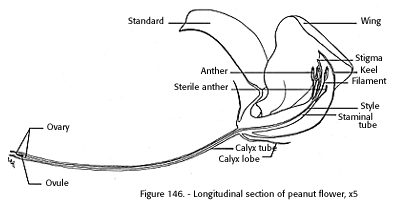
The first flowers appear near the base of each branch, 4 to 6 weeks after planting. Flowering continues along the branch for 6 weeks or more. The peanut flower is yellowish and about one-half inch in size. There may be one to several flowers in an inflorescence on the plant, but only one opens on one day, and there is an interval of one to several days between the opening of successive flowers. Smith (1950) stated that the peanut flower has a recurved beaked keel, with two petals fused along the dorsal edges to the apex but open ventrally at the base. There is a club-shaped stigma on a tortuous style extending beyond the eight functional and two sterile stamens (fig. 146).
The flower opens at sunup and pollen shedding occurs at once, the pollen accumulating between the anthers and stigma. Fertilization occurs 8 to 9 hours after pollination (Oakes 1958). After pollination, the flower fades (Beattie and Beattie 1943), and the ovary elongates to become the peg, which pushes into the soil (fig. 147). In 7 to 10 weeks, the peg matures into the reticulated pod of one to five edible seeds separated by slight constrictions (Gregory et al. 1951).
The value of peanut flowers to bees is not clear. Apparently, there is no functional nectary within the flower although some references indicate (erroneously) that bees collect peanut nectar (Graham 1941*, Pellett 1947*). Pollen is collected by honey bees although peanut plants are not considered to be a major pollen source by beekeepers.
[gfx] FIGURE 146.- Longitudinal section of peanut flower, x 5.
FIGURE 147.- Flower and pegs of a peanut plant.Pollination Requirements:
There is no doubt that peanuts are largely self-fertilized. The question is whether an increase in the set of seed is caused by cross-pollination. Some selections have a structure that impedes selfing and facilitates cross-pollination by bees. Reed (1924) reported that cross-pollination between cultivars occurs. Kushman and Beattie (1946) and Balhuis (1951) reported finding hybrids in peanuts. Stokes and Hull (1930) pointed out that the stigma of the mature flower " . . . usually lies buried among the dehisced anthers in the tightly closed keel petal so that self-fertilization is assured except for visitation by insects." Srinivasalu and Chandrasekaran (1958) noted that varietal differences exist for cross- pollination in relation to the protrusion of the stigma out of the keel. Leuck and Hammons (1969) reported that two cultivars have a structure in the flower that impedes self-pollination but which facilitates cross- pollination by bees. Leuck and Hammons (1965a) obtained no hybrids from caged plants but got a significant number from plants not caged. Later, they (1965b) reported that at least 80 percent of the peanut flowers in the open were actually tripped for pollen by bees.
Girardeau and Leuck (1967) showed that caged flowers not manipulated in any way produced 4 to 11 percent fewer fruits than hand flexed or water-drip manipulated flowers. Also, their open plots produced a significant 6 to 11 percent more than plots caged to exclude bees. Culp et al. (1968) recorded differences in the amount of crossing that occurred in different areas and in different cultivars but made no mention of the relation of pollinating insects to these differences. Shibuya et al. (1955) associated one-seededness with insufficient pollen on the stigma.
Pollinators:
There seems to be no doubt that the peanut flowe may be cross- pollinated and that crossing is primarily by bees. Hammons and Leuck (1966) showed that thrips are vectors of peanut pollen but that halictid bees were the principal visitors to peanut flowers at Tifton, Georgia. Hammons (1963), at the same location, noted that Lasioglossum, Megachile, Bombus, and Apis species worked peanut flowers. Hammons et al. (1963) and Leuck and Hammons (1969) added Anthidium and Melissodes species to the list of visitors but gave major credit for cross-pollination to species of halictids and megachilids. Diwan and Salvi (1965) stated that Apis cerana generally ignored peanut flowers, but Heide (1923) stated that the flowers were visited "actively and persistently" by A. cerana, and that A. cerana visited the flowers from 7 to 9 p.m. Gibbons and Tattersfield (1969) reported that A. m. adansonii, Nomia spp., and Megachile spp. visited the flowers in the Malawi area of Africa.
Leuck and Hammons (1965b) stated, "We conservatively estimate that in 1964, at least 80 percent of the peanut flowers were actually tripped for pollen each day by species of the combined bee complex." Unfortunately, they gave no indication of the bee population density, floral visitation, or bees per unit of flowers that provided this tripping. Hammons et al. (1963) noted that the halictids and megachilids were most abundant during the cool morning hours when most efficient pollination of peanuts occurs, whereas honey bee activity was spread over the day. No consideration was given to changing the degree of honey bee visitation by concentrating their numbers in the area.
If the 6 to 11 percent increase, which Girardeau and Leuck (1967) attributed to bee pollination, can be consistently obtained, it is of sufficient importance that consideration should be given to building up the bee population of large peanut plantings. This could be done by "saturation pollination" with honey bees if their use could be proven practical.
Pollination Recommendations and Practices:
None.
LITERATURE CITED:
BALHUIS, G. G.
1951. [NATURAL HYBRIDIZATION IN GROUNDNUTS.] Landbouwkundig. Tijdschrift [Wageningen] 63: 447-455. [In Dutch.]BEATTIE, W. R., and BEATTIE, J. H.
1943. PEANUT GROWING. U.S. Dept. Agr. Farmers' Bul. 1656, 31 pp.CULP, T. W., BAILEY, W. K., and HAMMONS, R. O.
1968. NATURAL HYBRIDIZATION OF PEANUTS. Crop Sci. 8: 109.DIWAN, V. V., and SALVI, S. R.
1965. SOME INTERESTING BEHAVIOURAL FEATURES OF APIS DORSATA FAB. Indian Bee Jour. 27(1): 52.GIBBONS, R. W., and TATTERSFIELD, J. R.
1969. OUT-CROSSING TRIALS WITH GROUNDNUTS (ARACHIS HYPOGAEA L.). Rhodesia Zambia Malawi Jour. Agr. Res. 7: 71-75.GIRARDEAU. J. H., and LEUCK. D. B.
1967. EFFECT OF MECHANICAL AND BEE TRIPPING ON YIELD OF THE PEANUT. Jour. Econ. Ent. 60: 1454-1455.GREGORY, W. C., SMITH, B. W., and YARBROUGH, J. A.
1951. MORPHOLOGY GENETICS AND BREEDING. In The PeanutÑthe Unpredictable Legume - a Symposium, pp. 2838, The National Fertilizer Assoc.HAMMONS, R. O.
1963. ARTIFICIAL CROSS-POLLINATION OF THE PEANUT WITH BEE-COLLECTED POLLEN. Crop Sci. 3: 562-563.______and LEUCK, D. B.
1966. NATURAL CROSS-POLLINATION OF THE PEANUT, ARACHIS HYPOGAEA L., IN THE PRESENCE OF BEES AND THRIPS. Agron. Jour. 58: 396.KROMBEIN, K. V., and LEUCK, D. B.
1963. SOME BEES (APOIDEA) ASSOCIATED WITH PEANUT FLOWERING. Jour. Econ. Ent. 56: 905.HEIDE, F. F. R.
1923. BIOLOGICAL OBSERVATIONS OF ARACHIS HYPOGAEA L. [Dutch East Indies] Alg. Proefsta. v. Landb. Meded. 14: 119.KUSHMAN, L. J., and BEATTIE, J. H.
1946. NATURAL HYBRIDIZATION IN PEANUTS. Amer. Soc. Agron. Jour. 38: 755.LEUCK, D. B., and HAMMONS, R. O.
1965a. FURTHER EVALUATION OF THE ROLE OF BEES IN NATURAL CROSS-POLLINATION OF THE PEANUT, ARACHIS HYPOGAEA L. Agron. Jour. 57: 94.______and HAMMONS, R. O.
1965b. POLLEN-COLLECTING ACTIVITIES OF BEES AMONG PEANUT FLOWERS. Jour. Econ. Ent. 58: 1028-1030.______and HAMMONS, R. O.
1969. OCCURRENCE OF TYPICAL FLOWERS AND SOME ASSOCIATED BEES (APOIDEA) IN THE PEANUT. ARACHIS HYPOGAEA. Agron. Jour. 61: 958-960.OAKES. A. J.
1958. POLLEN BEHAVIOR IN THE PEANUT (ARACHIS HYPOGAEA L.) Agron. Jour. 50: 387.REED, E. L.
1924. ANATOMY, EMBRYOLOGY AND ECOLOGY OF ARACHIS HYPOGAEA. Bot. Gaz.. 78: 289-310.SHIBUYA. T., SATO, H., and SUZUKI, M.
1955. [MORPHOLOGICAL AND PHYSIOLOGICAL STUDIES ON SUBTERRANEAN FRUCTUATION OF LEGUME CROPS.] Crop Sci. Soc. (Japan) Proc. 24: 16-19. [In Japanese, English summary.]SMITH B. W
1950. ARACHIS HYPOGAEA. AERIAL FLOWER AND SUBTERRANEAN FRUIT. Amer. Jour. Bot. 37: 802-815.SRINIVASALU. N., and CHANDRASEKARAN. N. R.
1958. A NOTE ON NATURAL CROSSING IN GROUNDNUT. ARACHIS HYPOGAEA LINN. Sci. Cult. [Calcutta] 23: 650.STOKES, W. E., and HULL, F. H.
1930. PEANUT BREEDING. Amer. Soc. Agron. Jour. 22: 1004-1019.