MUSKMELON
Cucumis melo L., family CucurbitaceaeThe muskmelons grown commercially in this country were classified by Whitaker (1970) into "varieties." Variety reticulatus Naud. includes the cantaloupes and 'Persian' melons, and variety inodorus Naud. includes the Casabas and the Honey Dews. There are numerous cultivars of each.
Muskmelons are grown in most States, but more than one half the acreage is in California. The bulk of the muskmelon crop is cantaloupes, combined with a small acreage of Casabas and 'Persians', which amounted 111,800 acres in 1970. Honey Dews were produced on 13,200 acres. The combined farm value of all muskmelons was $93.3 million.
Plant:
Muskmelons are trailing annuals, the vines, if unchecked, spreading to about 10 feet. The leaves are 4 to 8 inches across, and their 6- to 10- inch upright stem enables them to form a protective arborlike canopy over the flowers and fruit. The one to six melons per plant develop from the yellow hermaphrodite flower in the axis of the leaf. At maturity, about 6 weeks after the bloom appeared, the round to oblong melon is 4 to 8 inches in diameter. The frost-susceptible plants are usually grown in 6-foot rows, 4 to 24 inches apart in the row, with the best yield from plants 6 to 12 inches apart in the row (Pew 1952, Davis and Meinert 1966).
All forms of C. melo readily hybridize, as for example the 'Pershaw', which is thought to be a cross between the 'Persian' melon and the 'Crenshaw', Casaba, or the 'Honey Ball', which is a cross between the Honey Dew and the 'Texas Cannon Ball'. Rosa (1926) reported some self- incompatibility in the 'Persian' and the Honey Dew. The bulk of the discussion which follows will concern cantaloupes.
Inflorescence:
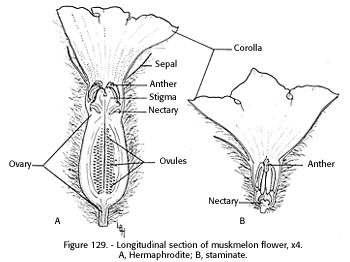
Most American cultivars of muskmelons are andromonoecious, bearing staminate and hermaphrodite flowers on the same plant (fig. 129). The numerous staminate flowers are borne in axillary clusters of three to five in all axillary positions not occupied by the few slightly larger solitary hermaphrodite flowers. The flowers are 3/4 inch to 1 1/2 inches across, with five petals united to slightly beyond the staminal column, then separated and broadly spreading (Whitaker and Davis 1962*). Griffin (1901) reported 512 staminate and 42 hermaphrodite cantaloupe flowers per vine. McGregor (1951 ) showed, however, that this ratio varies depending upon bee activity and fruit set. When bees were excluded, no fruit set and the ratio was one hermaphrodite to four staminate flowers, but in caged and open plots visited by bees the ratio was one hermaphrodite to 10 staminate flowers. Apparently, failure of the plant to set fruit stimulates production of a higher proportion of hermaphrodite flowers.
The staminate flower, supported on a thin stem, consists of the corolla, a single whorl of five stamens, only a few millimeters long, two pairs of which are united, with the anthers almost filling the small corolla tube. At the base of the corolla, a rudimentary style is surrounded by the nectaries (Judson 1935). The hermaphrodite flower has anthers and a broad, usually three-lobed stigma on a 1- to 2- mm style, the base of which is surrounded by the nectaries. The corolla of the hermaphrodite flower is on the end of the elongated ovary (Jones and Rosa 1928*, Judson 1949).
The muskmelon flower opens some time after sunup, the time depending upon the sunlight, temperature, and humidity. When the temperature is low, the humidity is high, or the day cloudy, opening is delayed. The flower closes permanently in the afternoon of the same day. Bee activity begins on the flower shortly after it opens, reaches a peak at about 11 a.m., and ceases about 5 p.m. (McGregor and Todd 1952*). At Davis, Calif., the flower opening and attraction for bees is an hour or so later in the day (Mann 1953).
The flower is attractive to bees for both pollen and nectar. Collection of pollen by bees usually ends before noon, but nectar collection continues into the late afternoon. Only about 1 percent as much nectar per acre is secreted by muskmelons as is secreted by alfalfa. Foster et al. (1965) showed that muskmelon plants infected with mosaic viruses produce less nectar than healthy plants.
[gfx] FIGURE 129. - Longitudinal section of muskmelon flower, x4. A, Hermahphrodite; B, staminate.
Pollination Requirements:
The isolation of muskmelon plants from pollinating insects and the caging of bees on the plants have proven that hermaphrodite flowers are incapable of performing self-pollination. The pollen must be transferred from the anthers to the stigma by insects (Alex 1957a, b; Bohn and Davis 1964; Mann 1953,1954; Mann and Robinson 1950; McGregor and Todd 1952 *,1952; and McGregor et al. 1965).
Muskmelons with fewer than 400 seeds are usually so small they are classed as culls. At least one viable pollen grain must be deposited on the stigma and fertilize an ovule if a seed is formed. The effective period in which this pollen can be deposited on the stigma is no more than a few hours in the morning, and if the temperature is high, the period may be only a few minutes. Single massive deposits of pollen by hand on the stigma are seldom as effective in the setting of fruit as repeated bee visits (Mann and Robinson 1950, Wolf and Hartman 1942). Muskmelon flowers are self-fertile (although not self-fertilizing), but when pollen comes from a different plant, the fruit that results may be slightly heavier (Rosa 1926). Also, a high correlation exists between the number of seeds in a muskmelon and its size - the more seeds the larger the fruit. Increased bee visitation is associated with greater number of seed.
Pollinators:
Tontz (1944) mentioned ants as possible pollinators of muskmelons and squash. Annand (1926) indicated that thrips might be pollinators of muskmelons, but Tsyganov (1953) considered one bee equal to 11,000 thrips. The value of thrips and ladybird beetles was discounted by McGregor and Todd (1952*, 1952) when they obtained no set of marketable cantaloupes in cages where honey bees were excluded and these insects were common, but a satisfactory set in cages supplied with bees. Bohn and Mann (1960) showed, with the mutant nectarless, the dependence of high muskmelon yields on honey bee pollination. The value of bees as pollinators of muskmelons, stated by Beattie and Doolittle (1926), Ivanoff (1947), Rosa (1927), and Rosa and Garthwaite (1926) is now firmly established. Because of their relative abundance in commercial fields and their attraction to muskmelon flowers, honey bees are the most important of the muskmelon pollinators. Beattie and Doolittle (1926) stressed the need for bees on muskmelons grown in greenhouses.
Honey bees visit muskmelon flowers as soon as the flowers open (fig. 130). They collect both nectar and pollen, move freely from flower to flower and plant to plant, and continue visiting the flowers until late afternoon. McGregor et al. (1965) showed that a honey bee visit to each flower about every 15 minutes is desirable for maximum fruit set. They calculated that one bee for each 10 hermaphrodite flowers is necessary to provide this rate of visitation. Whitaker and Bohn (1952) showed that variations in visits by honey bees occur between plants sometimes only a few feet apart if there is a variation in the microclimate around the plants. This means that many flowers must receive more visits than necessary if all are to receive the optimum number.
Growers prefer muskmelon fruit that is produced near the base of the plant. Such fruit is referred to as "crown set," or the set of fruit from the hermaphrodite flowers on the first to third spur. When there is heavy bee activity, a heavy crown set results (Rosa 1924, Whitner 1960). Such fruits are sweeter (McGregor and Todd 1952*, 1952), and are usually more oval than later fruits, which tend to be oblong.
Iselin et al. (1974) grew cantaloupes in an air-inflated plastic greenhouse. Their plants, shielded from bees, set no fruit, but plants visited by bees fruited normally. Bee foraging activity was similar to activity outside the greenhouse. They also reported that raising the CO2 content of the air in the enclosure increased the soluble solids (sugar content) of the ripe melons from 8 to about 12 percent.
Usually, the set of one or two fruits prohibits the set of further fruit until the first ones mature. Thus, when McGregor and Todd (1952) excluded bees for 3 weeks after initial flowering and then permitted unlimited visits to the flowers, 80 percent of the marketable fruit was set within the first 3 days, but the total production was not significantly different from production in cages where bees were constantly present. The fruits that set later were less sweet than crown-set fruit.
In studies on hybrid vigor in muskmelons, Foster (1963, 1967,1968a, b, c), Foster and Levin (1967), and Bohn and Davis (1957) found that F1 hybrids produced twice as much fruit as commercial cultivars, and other characters were improved. Bohn and Whitaker (1949) reported male sterility in the muskmelon, a character useful in hybrid seed production. Munger (1942) also showed that utilization of hybrid vigor was practical. In the utilization of hybrid vigor, pollination by bees is essential.
Taylor (1955) studied the production in 37 muskmelon fields in the Salt River Valley of Arizona in relation to proximity to honey bee colonies. In 20 fields with an average of one-half colony per acre within a mile, production was 1.06 melons per plant and 242 crates per acre. In 17 fields with no hives of bees in the "visible vicinity," production was only 0.67 melon per plant and 161 crates per acre. Honey bees were visiting muskmelon flowers in all fields.
[gfx] FIGURE 130. - Honey bee visiting muskmelon flower.
Pollination Recommendations and Practices:
Bees, primarily honey bees, are the major pollinating agents of muskmelons. The number of bees necessary for maximum pollination is the critical question. Taylor (1955) showed the economic significance of an inadequate supply. McGregor et al. (1965) demonstrated that one honey bee for each 10 hermaphrodite flowers should insure maximum pollination. This figure has not been extrapolated into colonies per acre - a rate that varies with conditions in, as well as beyond, the field. McGregor and Todd (1952*,1952) suggested one colony per acre for maximum muskmelon production. Peto (1951) used one to five colonies per acre on small fields. Pew et al. (1956) recommended one colony per acre placed in the shade on the edge of the field, but Eckert (1959*), without supporting data, recommended only one colony per 2 acres. Rupp (1969) reported a decrease of pollinated flowers with distance from the apiary (only 18 percent set on plants 600 m away but 40 percent set on plants within 100 m of the apiary), but he gave no indication as to the ratio of colonies per acre or bees per flower. Sims (1960) recommended one good strong colony per acre, the colony filling two deep hive bodies and having 750 to 1,000 in2 of brood. The Arizona Agricultural Experiment Station (1970) recommended one bee per 100 flowers in the field.
Because of the great increase in the number of flowers on the vine as the plant growth increases, the number of colonies required to provide this number might vary from a small fraction of a colony per acre to several colonies. Providing one honey bee for each 10 hermaphrodite flowers is the safest way to insure an adequate pollinator population at all times.
Practically all of the research on the pollination of C. melo has been on cantaloupes. The flower structure of the other types of muskmelons are identical or similar to that of cantaloupes. Until evidence is presented to the contrary, the assumption would appear to be safe that the pollination requirements are also the same for all cultivars of muskmelons.
LITERATURE CITED:
ALEX, A.H
1957a. HONEYBEES AID POLLINATION OF CUCUMBERS AND CANTALOUPES. Gleanings Bee Cult. 85: 398 - 400.______ 1957b. HONEY BEES AID POLLINATION OF CUCUMBERS AND CANTAL0UPS. Tex. Agr. Expt. Sta . Prog. Rpt. 1936,4 pp.
ANNAND, P. N.
1926. THYSONAPTERA AND THE P0LLINATION OF FL0WERS. Amer. Nat. 60: 177 - 182.ARIZONA AGRICULTURAL EXPERIMENT STATION.
1970. MELONS AND CUCUMBERS NEED BEES. Ariz. Agr. . Expt. Sta. and Ext. Serv. Folder 90.BEATTIE, J. H., and DOOLITTLE, S. P.
1926. MUSK-MEL0NS. U.S. Dept. Agr. Farmers' Bul. 1468, 38 pp.BOHN, G. W., and DAVIS, G. N.
1957. EARLINESS IN F1 HYBRID MUSK-MEL0NS AND THEIR PARENT VARIETIES. Hilgardia 26: 453 - 471.______and DAVIS, G. N.
1964. INSECT P0LLINATION IS NECESSARY FOR THE PRODUCTION OF MUSKMEL0NS (CUCUMIS MEL0 V. RETICULATUS). Jour. Apic. Res. 3(1): 61-63.______and MANN, L. K.
1960. NECTARLESS, A YIELD-REDUCING MUTANT CHARACTER IN THE MUSKMEL0N. Amer. Soc. Hort. Sci. Proc. 76: 455-459.______and WHITAKER, T. W.
1949. A GENE FOR MALE STERILITY IN THE MUSKMEL0N (CUCUMIS MEL0 L.). Amer. Soc. Hort. Sci. Proc. 53: 309 - 314.DAVIS G. N., and MEINERT, V. G. H.
1965. THE EFFECT OF PLANT SPACING AND FRUIT PRUNING ON THE FRUITS OF P.M.R. NO. 45 CANTAL0UPE. Amer. Soc. Hort. Sci. Proc. 87: 299 - 302. 260FOSTER, R. E.
1963. GLABROUS, A NEW SEEDLING MARKER IN MUSKMELONS. Jour. Hered. 54: 113-115.______ 1967. F1 HYBRID MUSKMELONS. I. SUPERIOR PERFORMANCE OF SELECTED HYBRIDS. Amer. Soc. Hort. Sci. Proc. 91: 390 - 395.
______ 1968a. F1 HYBRID MUSKMELONS. V. MONOECISM AND MALE STERILITY IN COMMERCIAL SEED PRODUCTION. Jour. Hered. 59(3): 205-207.
______ 1968b. F1 HYBRID MUSKMELONS. III. FIELD PRODUCTION OF HYBRID SEED. Amer. Soc. Hort. Sci. Proc. 92: 461 - 464.
______ 1968c. F1 HYBRID MUSKMELONS. IV. ROGUEING-THINNING TO PURE STANDS FROM MIXED SEED. Ariz. Acad. Sci. Jour. 5(4): 77-79.
______and LEVIN, M. D.
1967. F1 HYBRID MUSKMELONS. II. BEE ACTIVITY IN SEED FIELDS. Ariz. Acad. Sci. Jour. 4: 222 - 225.
______LEVIN, M. D., and MCGREGOR, S. E.
1965. NECTAR PRODUCTION BY MUSKMELONS INFECTED WITH FOUR MOSAIC VIRUSES. Amer. Soc. Hort. Sci. Proc. 86: 433-435.GRIFFIN, H. H.
1901. THE CANTALOUPE. Colo. Agr. Expt. Sta. Bul. 62, 18 pp.ISELIN, W. A., JENSEN, M. H., and SPANGLER, H. G.
1974. THE POLLINATION OF MELONS IN AIR INFLATED GREENHOUSES BY HONEY BEES. Environ. Ent. 3: 664-666.IVANOFF, S. S.
1947. NATURAL SELF-POLLINATION IN CANTALOUPS. Amer. Soc. Hort. Sci. Proc. 50: 314-316.JUDSON, J. E.
1935. THE FLORAL DEVELOPMENT OF THE STAMINATE FLOWER OF THE HONEY ROCK MUSK-MELON. West Va. Acad. Sci. Proc. 8: 93-98.______ 1949. THE FLORAL DEVELOPMENT OF THE PISTILLATE FLOWER OF CUCUMIS MELO. West Va. Acad. Sci. Proc. 20: 79-84.
MANN, L. K.
1953. HONEY BEE ACTIVITY IN RELATION TO POLLINATION AND FRUIT SET IN THE CANTALOUPE (CUCUMIS MELD). Amer. Jour. Bot. 40: 545-553.McGREGOR, S. E, LEVIN, M. D., and FOSTER, R. E.
1965. HONEY BEE VISITORS AND FRUIT SET OF CANTALOUPS. Jour. Econ. Ent. 58: 968-970.MUNGER, H. M.
1942. THE POSSIBLE UTILIZATION OF FIRST GENERATION MUSKMELON HYBRIDS AND AN IMPROVED METHOD OF HYBRIDIZATION. Amer. Soc. Hort. Sci. Proc. 40: 405 410.PETO, H. B.
1951. POLLINATION OF CUCUMBERS, WATERMELONS AND CANTALOUPES. In lowa State Apiarist Rpt. 1950. pp. 79-87.PEW, W. D.
1952. SIX-INCH SPACING UPS CANTALOUP YIELD. Prog. Agr. in Ariz. 3(4): 6 - 7.______MARLATT, R. B., and HOPKINS, L.
1956. GROWING CANTALOUPES IN ARIZONA. Ariz. Agr. Expt. Sta. Bul. 275, 24 pp.ROSA, J. T.
1924. FRUITING HABIT AND POLLINATION OF CANTALOUPE. Amer. Soc. Hort. Sci. Proc. 21: 51-57.______ 1926. DIRECT EFFECT OF POLLEN ON FRUIT AND SEEDS OF MELON. Amer. Soc. Hort. Sci. Proc. 23: 243 - 249.
______ 1927. RESULTS OF INBREEDING OF MELONS (CANTALOUPS). Amer. Soc. Hort. Sci. Proc. 23: 79-84.
______and GARTHWAITE, E. L.
1926. CANTALOUPE PRODUCTION IN CALIF. Calif. Agr. Expt.. Sta. Cir. 308, 48 pp.RUPP, K.
1969. INVESTIGATIONS ON POLLINATION ACTIVITY OF HONEYBEES. In 22d Internatl. Apic. Cong. Proc., Munich, pp. 557-560.SIMS, W. L.
1960. POLLINATION AND FRUIT SET IN CANTALOUPES. USE AND CARE OF HONEYBEES TO INSURE ADEQUATE POLLINATION. Calif. Coop. Ext. Serv. Vegetable Briefs - for Calif. Farm Advisors, 79, 4 pp.TAYLOR, E. A.
1955. CANTALOUP PRODUCTION INCREASED WITH HONEY BEES. Jour. Econ. Ent.. 48: 327.TONTZ, C.
1944. ANTS PINCH-HIT FOR BEES. Gleanings Bee Cult. 72: 482.TSYGANOV, S. K.
1953. [REMARKS ON THE POLLINATING ACTIVITIES OF HONEY BEES.] Uzbekistan Akad. Nauk. Inst. Zool. i parasitol. Trudy (1): 91 - 122. [ In Russian. ]WHITAKER, T. W.
1970. MUSKMELON VS. CANTALOUPE. HortScience 5(2): 86.______and BOHN, G. W.
1952. NATURAL CROSS POLLINATION IN MUSKMELON. Amer. Soc. Hort. Sci. Proc. 60: 391 - 396.______and DAVIS, G. N.
1962. CUCURBITS, BOTANY, CULTIVATION AND UTILIZATION. 250 pp. Leonard Hill (Books) Ltd. Interscience Publishers Inc., New York.WHITNER, B. F., JR.
1960. SEMINOLE - A HIGH-YIELDING, GOOD QUALITY, DOWNY AND POWDERY MILDEW-RESISTANT CANTALOUPE. Fla. Agr. Expt. Sta. Cir. S 122, 6 PP.WOLF, E. A., and HARTMAN, J. D.
1942. PLANT- AND FRUIT-PRUNING AS A MEANS OF INCREASING FRUIT SET IN MUSKMELON BREEDING. Amer. Soc. Hort. Sci. Proc. 40: 415 - 420.