GRAPES (INCLUDING RAISINS AND CURRANTS)
Vitis spp., family VitaceaeThe bulk of the grapes produced for the U.S. market are from the many cultivars of the Old World grape, or the "grape of history" (Vitis vinifera L.), grown on rootstock of American species resistant to the grape phylloxera insect (Snyder 1937). Production of table wine, and raisin grapes in California exceed by several times that of all of the other States combined. In 1969, California produced 3,600,000 tons of grapes on 457,266 acres. The leading cv. was 'Thompson Seedless' -a raisin, wine, or table grape- with 232,637 acres. Other important cvs. include other table grapes: 'Emperor' and 'Flame Tokay', and the wine grapes: 'Carignane', 'Grenoche', and 'Zinfandel'. The total production of grapes in all States was 3,902,510 tons, with a value of $273 million. The utilization of grapes was as follows: For wine and grape juice production, 2,258,757 tons; dried (raisins and currants), 1,015,200 tons; fresh, 557,179 tons; canned, 66,300 tons; and home use 5,084 tons.
Other species of grapes include the only native grape grown commercially, the muscadine grape (V. rotundifolia Michx.), the most important cv. being the 'Scuppernong', and the native bunch grape, which have been developed from one or more native species, sometimes hybridized with V. vinifera. These more important native species include V. aestivalis Michx., the summer or pigeon grape; V. labrusca L., the fox grape; V. Iincecumii Buckl., the post-oak grape; and V. vulpina L., the winter grape. Bailey (1949*) includes V. labruscana Bailey, which was derived from V. labrusca, and which is the source of the numerous cultivars grown commercially in the northeast. Hedrick (1924) stated that more than 2,000 cultivars of grapes are described in American viticultural literature and as many more in European literature.
Plant:
The grape is a climbing deciduous woody perennial, with 3- to 6- inch, heart-shaped leaves, inconspicuous panicled flowers, and a cluster of a few to 100 or more spherical or ovoid white, greenish, red, purple, or black fruit (a berry) l/4 inch to over 1 inch in size. In cultivation, the vines are frequently pruned and trained on trellises, 3 to 5 feet high, in such a way that the clusters of fruit can be harvested conveniently, some mechanically. Hundreds of cultivars are grown for different types of wine and grape juice production, other cultivars are grown for table grape use, or for drying as raisin or currant grapes.
Interplanting of cultivars is necessary for self-sterile cultivars, however, selection for increased degree of self-fertility has eliminated most of the self-sterility. Now most fields are solid plantings, sometimes scions from a single plant.
Inflorescence:
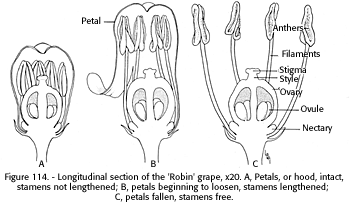
The grape flower cluster is a pyramid-like, loosely branched panicle, 1 to 10 inches long, containing up to several hundred inconspicuous greenish florets about one-quarter inch long. The floret usually has five stamens, but the number may range from two to seven (Randhawa and Sharma 1960), and five green petals (fig. 114). The stamens are about as long as the pistillate column. At the base of the ovary between the stamens are five, rarely six, yellow, fleshy nectaries. Insects are attracted to the flower by the nectar and pollen.
Kerner (1897*, p. 211) reported the unusual method of opening of the grape flower. The petals never separate at the top, but are united and serve as a domelike covering for the stamens and ovary. When these organs are mature and ready to function, the petals separate from the flower base, roll up spirally, and remain hanging together for a while like a hood which is finally thrown off by the tension in the expanding stamens. In midmorning of a warm calm day these caps fall like a gentle rain beneath the vine. Munson (1899) stated that there were three kinds of grape flowers - perfect, staminate, and pistillate.
Knuth (1908*, pp. 250-263) stated that V. vinifera flowers were complete, the stigma maturing simultaneously with the stamens but remaining receptive after the anthers have withered, making both self- and cross-pollination possible.
Pammel and King (1930*, pp. 1070 - 1072), stated that grape blossoms are visited by bees for nectar and pollen, and when the cap is released, pollen is thrown on the insect. Pellett (1947*) reported that the nectar yield, in terms of honey production to colonies of honey bees, is not great but that it is of some value. He considered the plant as a better source of pollen than nectar, with honeydew sometimes gathered from the leaves. Sharples et al. (1965) studied the pollination of the 'Cardinal' cv. of V. vinifera and concluded that bees were attracted to the flowers solely for pollen, with no functional nectaries present. Davydova (1969) also reported that honey bees visit grapes primarily for pollen.
[gfx] FIGURE 114. - Longitudinal section of the 'Robin' grape, x 20. A, Petals or hood, intact, stamens not lengthened; B, petals begining to loosen, stamens lengthened; C, petals fallen, stamens free.
Pollination Requirements:
The pollination requirements of grapes, somewhat like that of citrus (see "Citrus") are complex and for similar reasons; namely, different species, hybrids, and cultivars are involved. The pollination picture is further clouded by the fact that these have been intercrossed and selected for self-fertility, and that the observations have been reported over a long period of time, from different areas and at different stages in the development of a cultivar. Olmo (1936) spoke of one cultivar of V. vinifera that was parthenocarpic, being capable of producing fruit without pollination although its pollen was viable.
In general, V. vinifera has been considered self-fertile with the American species ranging from self-fertile to self-sterile (Beach 1892a, b, 1894, 1898; Booth 1911, Hedrick 1924). The muscadine (V. rotundifolia) is the most extensively grown example of a self-sterile species (Dearing 1938; Dickey and Loucks 1938; Husmanr 1916, 1932; Husmann and Dearing 1913, Reimer 1910; Reimer and Detjen 1910). Although Dearing (1917a, b) and Fry (1968) reported finding self-fertility in at least three muscadine selections, most cultivars now grown are self-sterile (Magoon and Snyder 1943). Other self sterile or partly self- sterile native American grape include 'Brighton', 'Herbert', and 'Salem' (Kell, 1944); 'Blue Lake' (Stover l960); and 'America', 'BarryÕ, 'Edna', 'Gaertner', 'Last Rose', 'Lindlye', 'Merrimac' and 'Munson' (Magoon and Snyder 1943).
Beach (1892a, b) thought that flowers with recurved stamens could not self but upright ones might self. Dorsey (1914) found 11 self-sterile or partly steril cultivars out of 95 with upright stamens examined, whereas only two of 37 cultivars with reflexed stamens were partly fertile. The others were sterile. This showed that although upright and recurved stamens were not positive proof that flowers were fertile or sterile, they indicated a likelihood - particularly of sterility - of those cultivars with reflexed stamens, with upright stamens being no surety of self-pollination.
In the case of the 'Ohanez' ('Almeria'), cv. of V. vinifera, there is no doubt about its self-sterility. Although pollen sprays have been used (Dunne 1942, Marriott 1950), the interplanting of other cultivars is considered better (Boehm 1960). Olmo (1943) was convinced that in at least some seasons honey bees were beneficial to this cultivar.
Probably the most thorough test of the pollination requirements of a V. vinifera cultivar was conducted by Sharples et al. (1965) on the 'Cardinal' cv. (Snyder and Harmon 1951). In this test, three mature plants were enclosed during the entire period of flowering in each of five 12- mesh-per-inch plastic screen cages containing a colony of honey bees. Five similar cages were used, which excluded all insects except those small enough to enter through the screen. The effect of pollinating insects on these plants was compared with that of five similar open plots.
The test revealed a correlation between seeds and berry weight as follows:
[gfx] fix table (spacing)
No. seeds per berry Mean weight per berry, grams 0 1.6 1 4.1 2 6.0 3 7.3 4 8.2 5 9.9
The berries in the bee cages and open plots had an average of 1.79 and 1.84 seeds each, compared to 1.65 seeds per berry in the no-bee cage. The clusters averaged 12.1 and 12.3 seedless berries in the bee cages and open plots compared to 16.1 seedless berries in the no-bee cage. This difference, though significant statistically, was not significant economically, in that the primary problem of eliminating shotberries (small, usually seedless berries) was not solved. A satisfactory crop was harvested from all vines. The effect of pollen from other grape cultivars or species was not determined but was proposed for future research.
Gladwin (1937) concluded that wind was responsible and that bees played only a minor role. Sharples et al. (1961) showed that clipping 1 cm from the inflorescence apex just before blooming was, alone, highly beneficial to quality of fruit set. Golodriga (1953) felt that different cultivars of V. vinifera reacted differently to pollinating agents.
There are other indications that even V. vinifera cultivars are benefited by insect pollination. The 'Ohanez' ('Almeria') cv. is noted for its self-sterility (Boehm 1960, Dunne 1942, Hale and Jones 1956, Magoon and Snyder 1943, Marriott 1950), but other cultivars also benefit from cross- pollination (Davydova 1969), Sosunov 1953, Steshenko 1958). Gladwin (1937) stated that cross-pollination is not only essential in self-sterile cultivars but that it is also beneficial in self-fertile cultivars. These benefits were substantiated by Iyer and Randhawa (1965), Laiok (1953), Lavrov (1956), and numerous others. Golodriga (1953) stressed the importance of selecting the proper pollenizer cultivars for those cultivars that shed or produce inferior berries. There seems to be no published information on the species and cultivars of grapes that have functional nectaries. The degree to which wind, compared to insects, is responsible for pollination and fruit set, has been established for only a handful of cultivars.
In general, modern grape specialists seem to have assumed that if some fruit set in a cluster in the absence of pollinating insects the plant was self-fertilizing, or wind pollinated, and any difference between "no special pollination problem" and "maximum production of quality fruit" was ignored.
The value of pollinating insects is given no consideration by growers of 'Thompson Seedless' grapes, yet it is well known among growers that cross-pollinated berries are long but self-pollinated ones are round and likely to shed. The degree to which insect pollinators might alter this relationship seems to have been given no consideration.
Pollinators:
There is also lack of agreement on the relative value of the pollinating agents on the cultivars of grapes known to benefit from cross- pollination. Einset (1930) insisted that insects cannot be depended upon. Gladwin (1937) gave major credit to wind and little credit to bees. Knuth (1908*, p. 250) and Munson (1899) considered both wind and insects of value, although the stigma is not adapted for wind pollination and the amount of pollen produced is small. Husmann and Dearing (1913) gave credit to a "small bee-like fly and a beetle," but later Husmann (1916) gave the credit to honey bees. Dearing (1938) considered the Halictus bee excellent but honey bees of sufficient value to warrant placing colonies in larger vineyards of V. rotundifolia and V. munsoniana J. H. Simson ex Planch. Reimer and Detjen (1910) and Olmo (1943) gave major credit to honey bees and flies, Steshenko (1958) to honey bees, Barskii (1956) reported that honey bees increased the weight of grape clusters by 23 to 54 percent, and Davydova (1969) associated pollinating insect visitation with increased yield and improved quality of grapes.
Laiok (1953) compared bee visitation and grape production of six cultivars in cages with and without bees. In five of the six cultivars, production was greater by 5 to 15 percent in cages with bees. Also, production in an open field decreased as distance from an apiary increased, with 220 kg from 10 bushes at the apiary, 180 kg at 200m, and 150 kg from 10 bushes at 600 m. On the other hand, Randhawa and Negi (1965) obtained no difference in set of open and self-pollinated plants of four cultivars.
Dearing (1938) considered Halictus bees excellent pollinators. Olmo (1943) gave considerable credit to the honey bee and a syrphid fly (Scaeva pyrastri (L.)). An across-the-board rating of insect visitors to grape flowers doubtless places the honey bee first.
Honey bees visit the flowers for pollen in the forenoon, primarily 9:30 to 11:30 a.m. (Sharples et al. 1965).
Some growers have objected to the presence of bees near their grapes under the mistaken belief that bees damage grapes, even though this claim has been disproved repeatedly for years (Clay 1886). If bees are rented for grape pollination they could easily be removed before the grapes are ripe because they do at times feed on the juice of grapes after the skin is broken.
Pollination Recommendations and Practices:
The recommendations for pollination of grapes for maximum production of highest quality fruit are not too consistent. In general, breeders have assumed that grapes were either completely self- fertilizing or were cross-pollinated by wind, so that in either case insects were considered of no value. Their assumption may be based in part on the construction of the flower, which would indicate that it is physically capable of transferring its pollen from the anthers to the stigma, or breeders may consider that a plant is self-fertile because bagged blossoms or isolated plants set fruit, without determining the maximum capability to set fruit.
In the case of most American species, and to an unknown degree the European (V. vinifera) species, there is evidence that insect visitation ranges from little or no value (Einset 1930, Sharples et al. 1965) to that of great benefit (Barskii 1956, Davydova 1969, Olmo 1943, Steshenko 1958). For example, there seems to be no doubt about the need for insect cross-pollination of the American species V. rotundifolia. Dearing (1938) recommended the placement of colonies of honey bees in larger muscadine vineyards for maximum production. Husmann (1916) also recommended the placement of colonies "here and there about the center" of muscadine vineyards of 100 acres or more. Reimer and Detjen (1910) recommended a hive of bees where "a large number of vines are maintained." Armstrong et al. (1934) and Armstrong (1935) recommended the interplanting of pollen- fertile cultivars within at least 50 feet of muscadine plants.
Steshenko (1958) stated that although grapes were normally wind- pollinated, bee visitation increased production. Sosunov (1953) and Shpakova (1961) agreed and recommended the interplanting of cultivars for maximum set. Davydova (1969) also agreed that grapes are wind- pollinated, but that bee visitation, mainly for pollen, increased yield and quality, so he recommended that one colony of honey bees be "appropriately located" per hectare. Barskii (1956) also recommended the use of honey bees but believed that one colony per 2 to 5 ha might be sufficient.
There are no recommendations for the use of bees on U.S. grapes. In most instances, cultivars are not inter-planted, and large vineyards are likely to be composed of scions of a single plant with no thought given to cross-pollination. The possible value of insect pollinators is given no consideration in grower recommendations. The evidence indicates that there may be a value, to some cultivars by such insect activity (Olmo 1943). In current agrotechnology, where the grower's net profit is a relatively low percentage of the gross income, even a minor increase becomes economically significant to him. With this thought in mind, a reappraisal of the significance of insect pollination of grapes seems to be justified.
LITERATURE CITED:
ARMSTRONG, W. D.1935. NEW VARIETIES AND POLLINATION OF MUSCADINE GRAPES. Amer. Soc. Hort. Sci. Proc. 33: 450-452. ______PICKETT, T. A., and MURPHY, M. M., JR. 1934. MUSCADINE GRAPES. Ga. Agr. Expt. Sta. Bul. 185, 29 pp. BARSKII, Y. A. S. 1956. [TRAINING BEES TO POLLINATE GRAPEVINES.] Sad i Ogorod (4): 64. [In Russian.] AA-391/61. BEACH, S. A. 1892a. THE SELF-POLLINATION OF THE GRAPE. Gard. and Forest 5: 451-452. ______ 1892b. NOTES ON SELF-POLLINATION OF THE GRAPE. N.Y. (Geneva) Agr. Expt. Sta. Ann. Rpt. 11: 597-606. ______ 1894. THE FERTILIZATION OF FLOWERS IN ORCHARDS AND VINEYARDS, ESPECIALLY IN ITS RELATION TO THE PRODUCTION OF FRUIT. N.Y. (Geneva) Agr. Expt. Sta. Ann. Rpt. 13: 633-648. ______ 1898. SELF-FERTILITY OF THE GRAPE. N Y. (Geneva) Agr. Expt. Sta. Bul. 157: 397 - 441. BOEM, E. W. 1960. SHOULD YOU POLLEN-SPRAY OHANEZ GRAPES? So. Austral. Dept. Agr. Jour. 64(5): 202-203. BOOTH, N. O. 1911. A SUGGESTION IN REGARD TO THE HISTORY OF GRAPE GROWING IN AMERICA. Amer. Soc. Hort. Sci. Proc., 18th Ann. Mtg.: 105Ñ 112. CLAY, O. M. 1886. A GRAPE-GROWER DEFENDING THE BEES. Amer. Bee Jour. 22: 469. DAVYDOVA, N. S. 1969. POSSIBILITIES OF EMPLOYING BEES TO POLLINATE VINEYARDS. In 22d Internatl. Apic. Cong. Proc., Munich, pp. 176-180. DEARING, C. 1917a. THE PRODUCTION OF SELF-FERTILE MUSCADINE GRAPES. Amer. Soc. Hort. Sci. Proc., 14th Ann. Mtg.: 30-34. ______ 1917b. MUSCADINE GRAPE BREEDING. Jour. Hered. 8: 409 - 424. ______ 1938. MUSCADINE GRAPES. U.S. Dept. Agr. Farmers' bul. 1785, 36 pp. DICKEY, R. D., and LOUCKS, K W. 1938. GRAPE GROWING IN FLORIDA. Fla Agr. Expt. Sta Bul. 324, 36 pp. DORSEY M. J. l914. POLLEN DEVELOPMENT IN VITIS WITH SPECIAL REFERENCE TO STERILITY. Minn. Agr. Expt. Sta. Bul.144, 60 pp. DUNNE, T. C. 1942. POLLEN-CONTAINING SPRAYS FOR THE CROSS-POLLINATION OF OHANEZ GRAPES. West. Austral. Dept. Agr. Jour. 19 (Ser. 2) (3): 210-213. EINSET, O. 1930. OPEN POLLINATION VS. HAND POLLINATION OF POLLEN-STERILE GRAPES. N.Y. (Cornell) Agr. Expt. Sta. Tech. Bul. 162, 14 pp. FRY, B. O. 1968. COWART, A NEW SELF-FERTILE MUSCADINE. Ga. Agr. Expt. Sta. Res. Rpt. 32, 6 pp. GLADWIN F. P. 1931. POLLINATION WITH PARTICULAR REFERENCE TO THE GRAPE. Amer. Fruit Grower 57(4): 16, 35. GOLODRIGA, P. I. 1953. [ON THE SELECTION OF POLLINIZER VARIETIES OF GRAPE.] Agrobiologiya 5: 105-110. [In Russian.] HALE, C. R., and JONES L. T. 1956. THE POLLINATION OF OHANEZ GRAPES. Jour. Agr. West. Austral. 5: 565 - 567. HEDRICK, U. P. 1924. MANUAL OF AMERICAN GRAPE-GROWING. 458 pp. The Macmillan Co., New York. HUSMANN, G. C. 1916. MUSCADINE GRAPES. U.S. Dept. Agr. Farmers' Bul. 709, 28 pp. ______ 1932. GRAPE DISTRICTS AND VARIETIES IN THE UNITED STATES U.S. Dept. Agr. Farmers' Bul. 1689, 33 pp. ______and DEARING, C. 1913. THE MUSCADINE GRAPES. U.s. Dept. Agr. Burl Plant Ind. Bul. 273, 64 pp. IYER, C. P. A., and RANDHAWA, G. S. 1965. HYBRIDIZATION STUDIES IN GRAPES: INVESTIGATIONS ON THE DIRECT INFLUENCE OF POLLEN ON SOME FRUIT AND SEED CHARACTERS. Indian Jour. Hort. 22(2): 107 - 121. KELLY, C. B. 1944. THE GRAPE IN ONTARIO. Ontario Dept. Agr. Statis. Branch Bul. 438, 38 pp. LAIOK V. D. 1953. [ON POLLINATION OF GRAPES BY HONEY BEES.] Pchelovodstvo 8: 46 - 48. [In Russian.] LAVROV, V. V. 1956. [INTERSPECIFIC POLLINATION OF GRAPES AND THE TRAINING OF BEES.] In Krishchunas, I. V., and Gubin, A. F., eds., Pollination of Agricultural Plants. MoskVa, Gos. Izd-vo. Selkhoz Lit-ry, pp. 200-203. [In Russian.] MAGOON, C. A., and SNYDER, E. 1943. GRAPES FOR DIFFERENT REGIONS. U.S. Dept. Agr. Farmers' Bul. 1936, 38 pp. MARRIOTT, P. F. 1950. POLLINATION OF TABLE GRAPES. Victoria Dept. Agr. Jour. 48(9): 391 - 394. MUNSON, T. V. 1899. INVESTIGATION IN AND IMPROVEMENT OF AMERICAN GRAPES Tex. Agr. Expt. Sta Bul. 56: 217-285. OLMO, H. P. 1936. POLLINATION AND THE SETTING OF FRUIT IN THE BLACK CORINTH GRAPE. Amer. Soc. Hort. Sci. Proc. 34: 402-404. ______ 1943. POLLINATION OF THE ALMERIA (OHANEZ) GRAPE. Amer. Soc. Hort. Sci. Proc. 42: 401-406. GRAPE 229 Literature Cited ARMSTRONG, W. D. 1935. NEW VARIETIES AND POLLINATION OF MUSCADINE GRAPES. Amer. Soc. Hort. Sci. Proc. 33: 450-452. ______PICKETT, T. A., and MURPHY, M. M., JR. 1934. MUSCADINE GRAPES. Ga. Agr. Expt. Sta. Bul. 185, 29 pp. BARSKII, Y. A. S. 1956. [TRAINING BEES TO POLLINATE GRAPEVINES.] Sad i Ogorod (4): 64. [In Russian.] AA-391/61. BEACH, S. A. 1892a. THE SELF-POLLINATION OF THE GRAPE. Gard. and Forest 5: 451-452. ______ 1892b. NOTES ON SELF-POLLINATION OF THE GRAPE. N.Y. (Geneva) Agr. Expt. Sta. Ann. Rpt. 11: 597-606. ______ 1894. THE FERTILIZATION OF FLOWERS IN ORCHARDS AND VINEYARDS, ESPECIALLY IN ITS RELATION TO THE PRODUCTION OF FRUIT. N.Y. (Geneva) Agr. Expt. Sta. Ann. Rpt. 13: 633-648. ______ 1898. SELF-FERTILITY OF THE GRAPE. N Y. (Geneva) Agr. Expt. Sta. Bul. 157: 397 - 441. BOEM, E. W. 1960. SHOULD YOU POLLEN-SPRAY OHANEZ GRAPES? So. Austral. Dept. Agr. Jour. 64(5): 202-203. BOOTH, N. O. 1911. A SUGGESTION IN REGARD TO THE HISTORY OF GRAPE GROWING IN AMERICA. Amer. Soc. Hort. Sci. Proc., 18th Ann. Mtg.: 105Ñ 112. CLAY, O. M. 1886. A GRAPE-GROWER DEFENDING THE BEES. Amer. Bee Jour. 22: 469. DAVYDOVA, N. S. 1969. POSSIBILITIES OF EMPLOYING BEES TO POLLINATE VINEYARDS. In 22d Internatl. Apic. Cong. Proc., Munich, pp. 176-180. DEARING, C. 1917a. THE PRODUCTION OF SELF-FERTILE MUSCADINE GRAPES. Amer. Soc. Hort. Sci. Proc., 14th Ann. Mtg.: 30-34. ______ 1917b. MUSCADINE GRAPE BREEDING. Jour. Hered. 8: 409 - 424. ______ 1938. MUSCADINE GRAPES. U.S. Dept. Agr. Farmers' bul. 1785, 36 pp. DICKEY, R. D., and LOUCKS, K W. 1938. GRAPE GROWING IN FLORIDA. Fla Agr. Expt. Sta Bul. 324, 36 pp. DORSEY M. J. l914. POLLEN DEVELOPMENT IN VITIS WITH SPECIAL REFERENCE TO STERILITY. Minn. Agr. Expt. Sta. Bul.144, 60 pp. DUNNE, T. C. 1942. POLLEN-CONTAINING SPRAYS FOR THE CROSS-POLLINATION OF OHANEZ GRAPES. West. Austral. Dept. Agr. Jour. 19 (Ser. 2) (3): 210-213. EINSET, O. 1930. OPEN POLLINATION VS. HAND POLLINATION OF POLLEN-STERILE GRAPES. N.Y. (Cornell) Agr. Expt. Sta. Tech. Bul. 162, 14 pp. FRY, B. O. 1968. COWART, A NEW SELF-FERTILE MUSCADINE. Ga. Agr. Expt. Sta. Res. Rpt. 32, 6 pp. GLADWIN F. P. 1931. POLLINATION WITH PARTICULAR REFERENCE TO THE GRAPE. Amer. Fruit Grower 57(4): 16, 35. GOLODRIGA, P. I. 1953. [ON THE SELECTION OF POLLINIZER VARIETIES OF GRAPE.] Agrobiologiya 5: 105-110. [In Russian.] HALE, C. R., and JONES L. T. 1956. THE POLLINATION OF OHANEZ GRAPES. Jour. Agr. West. Austral. 5: 565 - 567. HEDRICK, U. P. 1924. MANUAL OF AMERICAN GRAPE-GROWING. 458 pp. The Macmillan Co., New York. HUSMANN, G. C. 1916. MUSCADINE GRAPES. U.S. Dept. Agr. Farmers' Bul. 709, 28 pp. ______ 1932. GRAPE DISTRICTS AND VARIETIES IN THE UNITED STATES U.S. Dept. Agr. Farmers' Bul. 1689, 33 pp. ______and DEARING, C. 1913. THE MUSCADINE GRAPES. U.s. Dept. Agr. Burl Plant Ind. Bul. 273, 64 pp. IYER, C. P. A., and RANDHAWA, G. S. 1965. HYBRIDIZATION STUDIES IN GRAPES: INVESTIGATIONS ON THE DIRECT INFLUENCE OF POLLEN ON SOME FRUIT AND SEED CHARACTERS. Indian Jour. Hort. 22(2): 107 - 121. KELLY, C. B. 1944. THE GRAPE IN ONTARIO. Ontario Dept. Agr. Statis. Branch Bul. 438, 38 pp. LAIOK V. D. 1953. [ON POLLINATION OF GRAPES BY HONEY BEES.] Pchelovodstvo 8: 46 - 48. [In Russian.] LAVROV, V. V. 1956. [INTERSPECIFIC POLLINATION OF GRAPES AND THE TRAINING OF BEES.] In Krishchunas, I. V., and Gubin, A. F., eds., Pollination of Agricultural Plants. MoskVa, Gos. Izd-vo. Selkhoz Lit-ry, pp. 200-203. [In Russian.] MAGOON, C. A., and SNYDER, E. 1943. GRAPES FOR DIFFERENT REGIONS. U.S. Dept. Agr. Farmers' Bul. 1936, 38 pp. MARRIOTT, P. F. 1950. POLLINATION OF TABLE GRAPES. Victoria Dept. Agr. Jour. 48(9): 391 - 394. MUNSON, T. V. 1899. INVESTIGATION IN AND IMPROVEMENT OF AMERICAN GRAPES Tex. Agr. Expt. Sta Bul. 56: 217-285. OLMO, H. P. 1936. POLLINATION AND THE SETTING OF FRUIT IN THE BLACK CORINTH GRAPE. Amer. Soc. Hort. Sci. Proc. 34: 402-404. ______ 1943. POLLINATION OF THE ALMERIA (OHANEZ) GRAPE. Amer. Soc. Hort. Sci. Proc. 42: 401-406. 230 INSECT POLLINATION OF CULTIVATED CROP PLANTS RANDHAWA, G. S., and NEGI S. S. 1965. FURTHER STUDIES ON FLOWERING AND POLLINATION IN GRAPES. Indian Jour. Hort. 22(3/4): 287 - 308. ______and SHARMA, P. L. 1960. STUDIES ON FLOWERING AND POLLINATION IN GRAPES. Hort. Adv. 4: 21-37. REIMER, F. C. 1910. SELF-STERILITY OF ROTUNDIFOLIA GRAPES. Amer. Soc. Hort. Sci. Proc. 7th Ann. Mtg.: 27-32. ______and DETJEN, L. R. 1910. SEEF-STERILITY OF THE SCUPPERNONG AND OTHER MUSCADINE GRAPES. N.C. Agr. Expt. Sta. Bul. 209, 23 pp. SHARPLES, G. C., KUYKENDALL, J. R., TRUE, L. F., and TATE, H. F. 1961. IMPROVEMENT OF MARKET QUALITY OF CARDINAL GRAPE BY INFLORESCENCE APEX REMOVAL. Amer. Soc. Hort. Sci. Proc. 77: 316 - 321. TODD, F. E, McGREGOR, S. E., and MILNE, R. I. 1965. THE IMPORTANCE OF INSECTS IN THE POLLINATION AND FERTILIZATION OF THE CARDINAL GRAPE. Amer. Soc. Hort. Sci. Proc. 36: 321-325. SHPAKOVA, V. M. 1961. [BIOCHEMICAl ANALYSIS OF GRAPE POLLEN IN CONNECTION WITH SUPPLEMENTARY POLLINATION.] Sadov. Vinograd. i. Vinodel. (6): 26-28. [In Russian. ] AA-52/65. SNYDER, E. 1937. GRAPE DEVELOPMENT AND IMPROVEMENT. U.S. Dept. Agr. Yearbook 1937: 631-655. ______and HARMON' F. N. 1951. THE CARDINAL, CALMERIA AND BLACKROSE GRAPES FOR VINIFERA REGIONS. U.S. Dept. Agr. Cir. 882, 8 pp. SOSUNOV, V. I. 1953. [ON CROSSED POLLINATION OF GRAPES.] Sad i Ogorod 5: 26 - 27. [ln Russian.] STESHENKO, F. N. 1958. [THE ROLE OF HONEY BEES 1N CROSS-POLLINATION OF GRAPE VINES.] Pchelovodstvo 35: 37 - 40. [In Russian.] AA-182/60. STOVER, L. H. 1960. BLUE LAKEÑA NEW BUNCH GRAPE FOR FLORIDA HOME GARDENS. Fla. Agr. Expt. Sta. Cir. S-120, 10 pp.