EGGPLANT
Solanum melongena L., family SolanaceaeThe eggplant is a minor cooked vegetable crop in the United States. Florida with 2,350 acres and New Jersey with 1,400 acres in 1969 accounted for the bulk of the acreage, which was valued at $4,112,000. However, the plant is grown in home gardens in most areas of the country where there is a long, warm growing season. The average planting in Florida was 42 acres with a yield per acre of 565 bushels, for which the grower obtained an average of $1,310 per acre (Brooke 1970). The value of the 1969 crop in the U.S. was estimated by Brooke (1970) at $5.5 million.
Plant:
The eggplant is a much-branched, gray-green annual 20 to 50 inches high and appears somewhat like the pepper plant but is much coarser. The simple, thick, 6 to 15-inch leaves, are more or less oval, with the undemeath portion covered with thick, white woolly, sometimes spiny hairs. It is grown in rows and cultivated in a manner similar to that for peppers and tomatoes. The egg-shaped, purple fruit is usually harvested when near full size, 3 to 6 inches in diameter (fig. 107).
A fruit may have as many as 2,500 seeds (Odland and Noll 1948). Fruits are sometimes produced with few or no seeds, but they are hard and undesirable.
[gfx] FIGURE 107. - Eggplant, showing flower buds and fruit almost ready for harvest.
Inflorescence:
The 1 l/2 to 2-inch violet flowers of the eggplant are in two- or three- (rarely five) flowered cvmes. They may be perfect (Sambandam 1964) or hermaphrodite (Jones and Rosa 1928*). They develop opposite or near opposite the leaves instead of in the leaf axils as in most plants. The six to 20 anthers form a conelike tube around the style (fig. 108), and they dehisce at the terminal pores in a manner similar to that of the tomato flower, which favors self-pollination (Kakizaki 1924). However, the stigma ultimately projects beyond the anthers, where pollinating insects are more likely to contact it. This position affords ample opportunity for cross-pollination (Hawthorn and Pollard 1964*). The flower remains open 2 to 3 days without closing at night (Kakizaki 1924). It is visited by pollinating insects largely, if not exclusively, for pollen. Smith (1931) found that single flowers are less likely to shed than those on multiple cymes. Whether this is associated with pollination or some other factor is not clear.
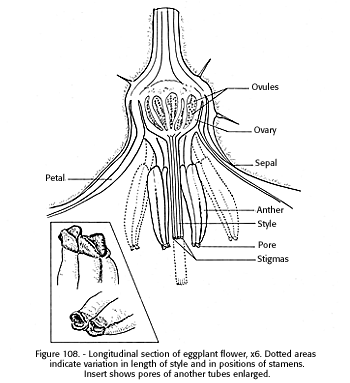
[gfx] FIGURE 108. - Longitudinal section of eggplant flower, x 6. Dotted areas indicate variation in length of style and in positions of stamens. Inset shows pores of anther tubes enlarged.
Pollination Requirements:
Bailey (1891) noted that artificial pollination always resulted in fewer seeds than natural pollination even when an excess of pollen was applied. He stated that with hand pollination a few seeds were produced at the apex of the fruit, but most of the ovules remained undeveloped. Jones and Rosa (1928*) reported that plants grown in a screened house isolated from insects were nonfruitful, and that flowers emasculated and left to natural pollination rarely set fruit. This indicated that the plant is not self-fruitful, that wind is not a factor in fruit set but that insects are required to transfer the pollen to the stigma in appropriate amounts and at the right time. The relative time period of pollen transfer for most effective fertilization of a flower has not been determined.
Jasmin (1964) reported that male-sterile plants have been found, in which the anthers do not dehisce. Such plants must be insect pollinated and might be used in the production of hybrid plants. Capinpin and Alviar (1949) reported that hybrids fruited earlier than the parents. Baha-Eldin et al. (1968) concluded that hybrid vigor was strongly manifested in total yield and number of fruit per plant, which would justify the utilization of heterosis in eggplant. Kakizaki (1931) reported that in most of his crosses the first harvesttime was earlier, and production exceeded the best parent by 17 percent. Hybrid eggplants are now being produced commercially by the use of this male sterility factor.
Pollinators:
Wind is not a factor in eggplant pollination, and vibration of the blossom will not cause a sufficient deposit of pollen on the stigma. The eggplant does not self without the aid of bees or man (Kakizaki 1924). The pollinating insects on eggplant have never been studied. Pammel and King (p. 606,1930*) reported that bumble bees were common on the flowers at Ames, Iowa, but no honey bees came to the flowers. Workers dealing with this crop have tended to overlook the insect visitors, but the amount of crossing recorded by different ones indicates that insect visitation occurs in relative abundance. Sambandam (1964), for example, stated that 30 to 40 percent of the fruit set is attributed to pollination by contact, gravity, and wind, the rest to insects, and he reported that crossing on the same plant (in India) ranged from 0.7 to 15 percent, but he made no mention of the insect pollinators responsible for the set or crossing. Kakizaki (1924) reported 0.2 to 46.8 percent cross-pollination. Pal and Taller (1969) likewise discussed pollination of eggplant, and stated that within the variety the number of seeds per fruit is higher in cross-pollinated than in selfed plants, but substantially lower than in open-pollinated plants. No mention is made of the pollinating insects responsible for the better effect on the open-pollinated flowers.
Kakizaki (1924) concluded that bees or man are necessary in the pollination of eggplants.
Pollination Recommendations and Practices:
If male-sterile plants are grown for the production of hybrids, the pollinating insects are essential and should probably be present in relatively large quantities. Even if fertile varieties are grown for fruit production, the meager evidence available strongly indicates that a goodly supply of pollinating insects should be available in the field.
LITERATURE CITED:
BAHA-ELDIN. S. A., BLACKHURST, H. T., and PERRY. B. A.
1968. THE INHERITANCE OF CERTAIN QUANTITATIVE CHARACTERS IN EGGPLANT (SOLANUM MELONGENA L.). H. INHERITANCE OF YIELD, FRUIT NUMBER AND FRUIT WEIGHT. Amer. Soc. Hort. Sci. Proc. 92: 490-497.BAILEY. L. H.
1891. EXPERIENCES WITH EGGPLANTS. N.Y. (Ithaca) Agr. Expt. Sta. Bul. 26, 26 pp.BROOKE, D. L.
1970. COSTS AND RETURNS FROM VEGETABLE CROPS IN FLORIDA SEASON 1968-69 WITH COMPARISONS. Fla. Agr. Expt. Sta. Agr. Econ. Rpt. 2, 34 pp.CAPINPIN. J. M., and ALVIAR, M. A.
1949. HETEROSIS IN EGGPLANT. Philippine Agr. 33: 126 - 141.JASMIN, J. J.
1954. MALE STERILITY IN SOLANUM MELONGENA L.: PRELIMINARY REPORT ON A FUNCTIONAL TYPE OF MALE STERILITY IN EGGPLANTS. Amer. Soc. Hort. Sci. Proc. 63: 443 - 446.KAKIZAKI, Y.
1924. THE FLOWERING HABIT AND NATURAL CROSSING IN THE EGG- PLANT. Japan Jour. Genet. 3: 29 - 38.____ 1931. HYBRID VIGOR IN EGG-PLANT AND ITS PRACTICAL UTILIZATION. Genetics 16: 1-25.
ODLAND, M. L., and NOLL. C. J.
1948. HYBRID VIGOR AND COMBINING ABILITY IN EGGPLANTS. Amer. Soc. Hort. Sci. Proc. 51: 417 - 422. PAL, G., andTALLER. M.
1969. [EFFECTS OF POLLINATION METHODS ON FERTILIZATION IN EGGPLANT (SOLANUM MELONGENA L.).] Acta Agron. Acad. Sci. Hung. 18(3/4): 307-315. [In Hungarian.] Abstract in Biol. Abs. 52(11) 58979: 5925. 1971.SAMBANDAM, C. N.
1964. NATURAL CROSS-POLLINATION IN EGGPLANT (SOLANUM MELONGENA). Econ. Bot. 18(2): 128 - 131.SMITH, O.
1931. CHARACTERISTICS ASSOCIATED WITH ABORTION AND INTERSEXUAL FLOWERS IN THE EGGPLANT. Jour. Agr. Res. 43(1): 83-94.