COTTON
Gossypium spp., family MalvaceaeCotton is grown primarily for its lint, although the seed, valued at about one-fifth that of the lint, is also used for planting or is crushed, a food oil recovered, and the residue used as a livestock food.
Cotton was naturally dispersed throughout much of the warmer parts of the world even before 3,000 B.C., when its lint was spun and woven into cloth (Gulati and Turner 1928). Only four of the numerous species of Gossypium are cultivated for their spinnable fibers. These are the two diploid Asiatic species, G. arboreum L. and G. herbaceum L., and the two amphidiploid species, G. barbadense L. and G. hirsutum L. The first two are confined primarily to the Old World. The last two are grown both in the New World and abroad. G. hirsutum, known as upland or short staple cotton, is grown most extensively. G. barbadense, known in the United States as American Pima, or Pima, and sometimes called extra-long staple cotton, is grown in long-season areas, such as our Southwestern States.
The crop is grown from Virginia southward and westward to California, in an area often referred to as the Cotton Belt. The more important cultivars 24 of upland cotton planted in 1971 included 'Deltapine 45', which accounted for 25 percent of the total acreage, 'Stoneville 213', which accounted for 18 percent, and 'Acala SJ-1', which accounted for 10 percent. Other major cultivars and their percentages included: 'Lankart LX 571', 7; 'Coker 201', 4; and 'Paymaster 111', 'Stoneville 7A', and 'Stripper 31', each 3 percent (USDA 1971).
Texas, California, Mississippi, and Arkansas were the leading cotton producing States in 1970. The value of the entire U.S. crop was approximately $1.2 billion. The leading cotton producing countries of the world in 1970 are shown in table 12.
The lint percentage of cotton varies from 30 to 40 percent, more often in the high 30's for upland and the low 30's for Pima.
[gfx] fix table 12
TABLE 12. - Leading cotton producing countries of the world in 19701 __________________________________________________________ Millions Pounds Millions of of lint of bales Country acres per acre of lint 2 ______________________________________________________________________________ India.................................................................... 19.0 111 4.4 China (mainland)............................................ 12.5 268 7.0 United States.................................................. 311.2 437 10.3 Russia................................................................. 6.8 761 10.8 Brazil.................................................................. 6.0 183 2.3 Pakistan............................................................ 4.3 271 2.4 United Arab Republic................................... 1.7 664 2.3 Turkey................................................................ 1.3 676 1.8 Sudan.................................................................. 1.3 406 1.1 Mexico................................................................ 1.0 692 1.4 __________________________________________________________ l Source: Anonymous 1972. 2 An average bale weighs 480 pounds. 3 Includes about 77,600 acres of Pima.Recognizing the variation in cotton due to natural speciation, breeder manipulation, the wide distribution, and the conditions under which it is grown, the following discussion will be largely confined to upland cotton. Pima and, to a much lesser degree, the other two species, will also be mentioned. Because of some lack of agreement on the pollination requirements of cotton, more than the usual amount of space is devoted herein to this crop.
___________
24 In cotton, a cultivar, or "variety," is neither a clone, a pure line, nor a primary mixture of pure lines. It is usually a progeny row selection, bulked and mass multiplied, during which time insect pollination may have played a major or insignificant role, depending upon the pollinator population present during the flowering season.Plant:
The cotton plant is a broad-leaved perennial, 2 to 5 feet tall, that is treated as an annual under much of its growing conditions in the United States. It becomes a perennial if the ground in which it grows does not freeze during the winter. The plants will grow and be productive on a wide variety of soils. It is most productive on fertile soil under hot weather and irrigated conditions if rainfall is deficient. The seeds are usually planted 4 to 8 inches apart in about 3-foot rows after all danger of frost is past in the spring. Flowering on the first of its lower branches begins in about 2 months and may continue on succeeding branches and growth another 2 months at about which time the first ripe fruit (bolls) begin to open and expose the mature lint-covered seed (fig. 87). Most of this raw cotton is currently machine-harvested then transported to a cotton gin where the lint is separated from the seed then pressed into bales.
Flowering and fruiting on the plant follows a spiral course from the innermost bud on the oldest and lowest fruiting branch, and ends on the latest growth toward the tip of the plant.
Pima cotton is usually more robust than upland cotton, with waxy- green leaves and smaller bolls. Fruiting on the plant begins later in the season, which tends to restrict its culture to the area with the longest growing season, such as the extreme southwestern United States.
Only about half as much lint per acre is normally produced on Pima cotton as is produced on upland cotton; however, the grower receives about twice as much for the more desirable lint, so the net profit from the two types of cotton is similar. The lint can be removed from the seed with the same type of gin saws that are used on upland cotton, but the quality of the lint is better if it is removed with a roller gin.
[gfx]
FIGURE 87. - Open bolls of cotton.Inflorescence:
The 2- to 4-inch-long by 2-inch-broad cotton blossom is subtended by three green leafy bracts, each an inch or more across, and a green calyx that fits snugly around the base of the ovary. The five-petal corolla of upland cotton is cream colored when it opens in the morning shortly after sunup, but turns pink in the afternoon and closes toward nightfall never to reopen (fig. 88). On the second day, the color of the petals is a watermelon-red. The typical corolla of Pima cotton is yellow, with a maroon throat or petal spot, and the color changes little with age. The corolla and stamina column usually fall on the second day.
The staminal column surrounds the 1- to 2-inch-long style leading from the ovary and terminating in the l/4 to l/2-inch-long stigma (fig. 89). The ovary contains 5 to 10 ovules in each of three to five sections, carpers, or locules. The stamina sheath, enclosing most of the style, bears numerous stamens l/4 to 1/2 inch long, each terminating in an anther that normally produces an abundance of viable self-fertile pollen, 45,000 grains per flower (Tsyganov 1953). The grains are large, 81 to 143 microns (Kaziev 1964), and coated with a viscid material that causes them to adhere to each other; therefore, cotton pollen is not transported by wind. Each section of the oval, 1-inch boll that develops from the ovary may produce a "lock," a distinct group of lint-entangled seed. These locks are exposed in the open three- to five-sectioned "burr."
The number of flowers on a cotton plant are determined by numerous factors including the available plant food, water supply, variety, and density of the plant population. Usually, about half of the flowers produce mature bolls (Brown 1938, Buie 1928, Dunlap 1945, McNamara et al. 1940, Sen and Afzal 1937). Flowering reaches its peak at about four flowers per plant per day. Between 225 and 400 bolls are usually required to produce a pound of fruit.
[gfx]
FIGURE 88. - Cotton flower, showing general corolla shape an proximity of anthers to stigma. A, Upland cotton flower with flared, cream-colored corolla, and stigma protruding only slightly above the anthers; B, Pima cotton flower, with tubular shaped yellowish corolla and dark "petal spot" toward base. Stigma extends well above the anthers.FIGURE 89. -Flower of 'Deltapine' upland cotton, x 1.5. A, Side view, showing one of the 3 subbracteal nectaries; b, bracts slightly spread to show one of the 3 inner bracteal nectaries; C, longitudinal section.
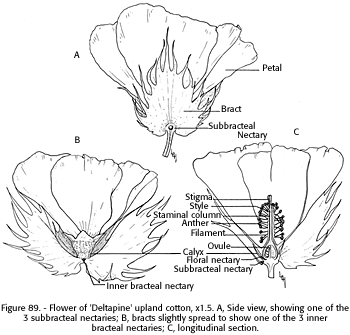
NECTARIES AND NECTAR SECRETION OF COTTON:
Nectar is normally produced in five different areas on the cotton plant, although the reason why the nectar is secreted is not clear. Trelease (1879) made a detailed report on cotton nectar secretion and its possible purposes (although he stated "Glover, Agricultural Report 1855, p. 234, mentions these glands - and their secretion of a sweet substance, which ants, bees, wasps, and plant bugs avail themselves of as food"). Kaziev (1964) stated that Delpino, in 1900, was the first to characterize the floral and foliar nectaries of cotton.
The different areas of nectar secretion are (1) floral, (2) inner or circumbracteal, (3) outer or subbracteal, (4) foliar or leaf, and (5) unipapillate (microscopic) areas on the flower peduncles and young leaf petioles (Mound 1962) (fig. 90). These unipapillate nectaries are rarely visited by pollinating insects, contribute nothing to pollination and little or nothing to the welfare of pollinating agents, and will not be further discussed. Trelease (1879) believed that the floral nectaries were associated with pollination but that extra-floral nectaries were associated with attracting harmful insects away from the delicate flower parts. Kottur (1921 ) believed that the nectar and pollen in the flower invited natural crossing.
In addition to the secretion from the nectaries, there is a saccharine exudation of certain aphids, white flies, and thrips on cotton, known in the Sudan as "asal" (Bedford 1921). When this material is present in abundance, it supports growth of a sooty fungus, causing a detrimental blackening of the cotton leaves. At times, the sticky material becomes mixed with the lint with damaging effects (Hadwich 1961). When honey bees are present in sufficient numbers, they collect this material as food, and by removing it they become beneficial in a sense other than as pollinators.
[gfx]
FIGURE 90. - Nectaries of cotton plant. A, Honey bee collecting nectar from a subbracteal nectary; B, inner bracteal nectaries; C, section of calyx removed to show proboscis (tongue) of bee (while bee is inside flower) reaching for floral nectar droplets; D, underside of leaf, showing location of leaf nectaries.FLORAL NECTARIES:
Within the flower, the nectar exudes from a ring of papilliform cells at the base of the inner side of the calyx (Tyler 1908). Secretion may begin a few hours to a few days before the flower opens, but, because of its unavailability to pollinators until the flower opens, this possible early secretion is of no consequence. The nectar reaches its maximum accumulation by mid-afternoon, the amount depending upon climatic factors, soil fertility, water, and cultivar involved (Kaziev 1959a, b, 1967), and ceases when the petal color begins to change, an indication, according to Kaziev (1964) and Mel'nichenko (1963) that pollination has occurred.
NECTAR COMPOSITION IN RELATION TO ATTRACTIVENESS TO BEES:
At times, honey bees appear to be noticeably reluctant to visit cotton blossoms, even though much nectar and pollen are present. Wykes (1952) studied the preference of honey bees for solutions of various sugars and found that preferences were shown for solutions of single sugars in the following descending order: Sucrose, glucose, maltose, and fructose, and that mixtures of equal parts of all these sugars was the most attractive combination. Vansell (1944a, b) studied the composition of sugars in orange and cotton floral nectar and found the percentages shown in table 13.
[gfx] fix table 13
TABLE 13. - Percentage of sugars and moisture content of cotton and orange nectars _________________________________________________________ Plant Levulose Dextrose Sucrose Approximate Moisture __________________________________________________________ Cotton: ÔAcalaÕ 14.27 13.06 0.71 70 ÔPimaÕ 10.36 9.25 .35 80 Orange: ÔWashington NavelÕ 6.46 5.42 12.87 75 ÔValenciaÕ 6.08 5.06 12.38 77 __________________________________________________________
Butler et al. (1972) collected nectar from leaf, floral, and extra- floral nectaries of 'Hopicala', 'Deltapine Smooth-leaf', 'Deltapine 16', and 'Pima S-2' near Tucson, Ariz., and analyzed it by gas chromatography for its sugars. They also found low sucrose percentages in nectar from floral and subbracteal nectaries.
Ivanova-Paroiskaya (1950) showed the fructose - glucose - sucrose percentages of floral nectar of G. barbadense cotton, cv. '35-1', to be 39.78-37.50-1.63, and for upland cotton, cv. '36 - 7 M', to be 37.85-35.65- 6.89. Kaziev (1964) showed that the range of the sucrose content of cotton nectars was from 2.3 to 7.6 percent, with the total mono-sugars ranging from 21.2 to 46.9 percent. All samples were taken during mid-season flowering. Whether the percentages change with the season has never been determined.
Numerous observations have shown a relatively low percentage of honey bee visits to flowers of cotton during mid-season and a high percentage toward the beginning and the end of the season. For example in August 1952 (unpublished data), at Tucson, Ariz., eight blossoms of 'Pima S-1' were observed constantly from 8:45 until 11:30 a.m. During that time, they received visits from one honey bee, one Bombus spp., 100 Melissodes spp., and five unidentified pollen-collecting bees. In the same plot, on October 10 between 7 a.m. and noon, three blossoms of 'Pima S-1' were visited by 363 honey bees and seven Melissodes bees. The reason for this extreme difference in the number of bee visitors is unknown.
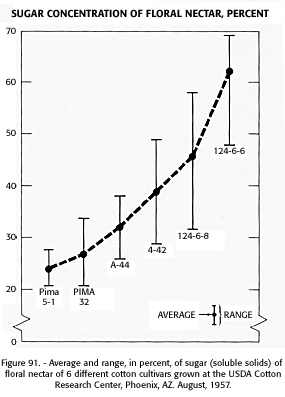
The volume of nectar in the blossoms of Pima cotton is greater than that in upland cotton, but the sugar concentration is lower. As much as 0.1 ml of nectar has been collected at one time from the former but never more than half this amount from upland. The sugar concentration of upland cotton floral nectar is greater (reaching a maximum of about 69 percent) than that of Pima nectar (a maximum of only 34 percent (fig. 91). The volume of floral nectar of both cottons exceeds the volume of extrafloral nectar, but, as previously stated, the floral nectar is less attractive to honey bees.
Nectar secretion of cotton is strongly influenced by soil fertility, as various tests made in Russia have shown. For example, superphosphate increased nectar secretion by 170 percent and potassium by 130 percent (Monokova and Chebotnikova 1955). Extensive studies by Kaziev (1964) showed that nitrogen had no effect on nectar production, but the greatest increases resulted from application to the soil of cattle manure alone or with complete fertilizers.
[gfx] fix diagram
FIGURE 91. - Average and range, in percent, of sugar (soluble solids) of floral nectar of 6 different cotton cultivars grown at the USDA Cotton Research Center, Phoenix, Ariz., August 1957.SUGAR CONCENTRATION OF FLORAL NECTAR, PERCENT 7n . Approximate _ _ Plant Levulose Dextrose Sucrose Moisture Cotton: 'Acala' 14.27 13.06 0.71 70 ~ , 'Pima' 10.36 9.25 .35 80 60 _ Orange: _ t 'Washington Navel' 6.46 5.42 12.87 75 'Valencia' 6.08 5.06 12.38 77 50 _ ~ _ _ ~_ _
NECTARIES OUTSIDE THE COROLLA (INVOLUCRAL NECTARIES):
There are usually three nectaries just below the sepals at the union of the three bracts, and three others just below the base of the bracts. These have variously been referred to as calyceal and subbracteal, inner and outer involucral, extrafloral internal and extrafloral external, and bracteal nectaries. They begin to function several days before the flower opens, but the day before opening they secrete nectar in copious amounts and this secretion may then continue from several days to 2 to 3 weeks after flowering. When beekeepers report good honey flows from cotton, the bees are usually working these involucral nectaries more intensively than the other nectaries of cotton. Kaziev (1964) also showed that these involucral nectaries were highly attractive to bees. Some upland cotton plants may not have all of these nectaries.
In Hawaiian cotton (G. tomentosum Nutt. ex Seem.), the nectaries are entirely absent. This characteristic has been transferred to some commercial selections to provide a nectariless cotton (Meyer and Meyer 1961). These selections are studied to determine the effect of nectar deprivation on harmful lepidopterious insects on cotton (Lukefahr and Martin 1964, Lukefahr and Rhyne 1960). The nectariless characteristic has been suggested in connection with proposed production of hybrid cotton, the theory being that if only floral nectar were present the bees would visit the flowers more frequently. The opposite, however, may be true, that is, the bees may be attracted to the field by the extrafloral nectaries and then discover the floral ones. Research in this area is requisite for a successful hybrid cotton program if pollinating insects are used.
FOLIAR OR LEAF NECTARIES:
Nectar is also produced on the underside of the leaf in usually one nectary on the primary vein near the blade and petiole juncture. Occasionally, there are two additional nectaries, one on a vein on either side of the primary vein. The leaf nectary begins to function before the leaf reaches full size and may function for 2 or 3 weeks. Leaf nectar secretion on the plant begins when the first flower is in the early bud, or "square", stage and may continue as long as the plant is producing new leaves (Kaziev 1964). The structure of cotton leaf nectaries was described by Reed (191 7).
Honey bees show preference for all of the extrafloral nectaries over floral nectar but under certain conditions collect nectar from all. Ivanova-Paroiskaya (1956) showed the following relative number of honey bee visits to the nectaries: Floral 32, calycular 219, subbracteal 580, and leaf 389. Many other insect species show a preference for the floral nectary of cotton.
COTTON AS A HONEY PLANT:
In many of the areas where cotton is grown it is considered a major honey plant (Benson 1937, Kuliev 19S8, Minkov 1957, Parks 1921). It does not produce as much honey per acre per day as alfalfa, clover, or many other sources (Butler et al. 1972) but because of its longer flowering period, excellent crops of honey can be obtained. Pima cotton is recognized by beekeepers as a better honey source than upland cotton. Vansell (1944a) calculated that 1 acre of Pima cotton was equal to 30 acres of Acala in the production of nectar. Ivanova-Paroiskaya (1950) reported honey crops of 300 kg/ha (267 lb/acre) for G. barbadense compared to 75 to 90 kg/ha (66 to 80 lb/acre) for upland. Normally, when bees are working cotton blossoms, there is a steady, but not spectacular, storage of the high-quality cotton honey. Unfortunately, highly toxic insecticides, frequently used on cotton during the flowering period, kill many bees and prevent the harvest of a honey crop.
Pollination Requirements:
Cotton is usually referred to as a partially cross-pollinated crop, although many breeders have treated it as a completely self-fertile and self-pollinating crop except for accidental and unwanted cross- pollination caused by pollinating insects. Cross-pollination has been referred to as "natural crossing," and is considered detrimental because of the introduction of off-type plants into the progeny. Its impact on production has not been given much consideration. Breeders know that a cotton blossom isolated by any one of several techniques will usually develop a mature boll with viable seed. Breeders also know that they seldom obtain by this method quite as many seed or as much lint from such bolls as they obtain from open-pollinated ones (Stephens 1956). Because this difference can be altered to the benefit of the grower by the activity of pollinating insects, and because of the association of pollinating insects with natural crossing and resultant undesirable cotton types, the subject of natural crossing is reviewed.
NATURAL CROSSING:
The term "natural crossing" is freely used in cotton literature, but not always precisely defined. The generally accepted meaning appears to be "the amount of cross-pollination effected by insects, as opposed to hand cross-pollination, that can be detected in breeding lines." Fryxell (1957) defined natural crossing as "that which occurs between individuals within a population." He stated that the two phenomena (intra- and inter- population crossing) are related, but the distinction is not always made clear. Simpson (1954a) stated that cross-pollination is not readily detected in cotton unless distinctive marker characters are present in the parental lines. For this reason, the extent of natural crossing in open- pollinated fields has been consistently underestimated by some cotton breeders. Natural crossing is usually associated with insect pollination of plants but seldom with its benefit to the cotton plant.
Various breeders have reported the percentage of natural crossing in their area and commented upon its detrimental effect. Balls (1912) reported 13.3 percent natural crossing of cotton in Egypt, and proposed the isolation of plants under mosquito netting to exclude bees. He noted, however, that some strains "resent this treatment and refuse to hold their bolls," a possible indication that he might have been unknowingly dealing with a degree of self-sterility in the plants, although shading of the plants might also have contributed.
Webber (1903) also reported 5 to 10 percent natural crossing between varieties of upland cotton grown in adjacent rows in the United States and concluded that absolute prevention of crossing would require isolation of cotton by 5 to 10 miles from other cotton. Ricks and Brown (1916) reported 2.8 to 18.5 percent in alternate hills in alternate rows. The type or preponderance of insect pollinators responsible for this cross- pollination was not given.
Cook (1921) stated that natural cross-fertilization in the field is one of the major causes of "running out" of varieties. Later, Cook (1932) stated that maintaining the seed stock of superior strains rather than creation of new ones was the essential breeding problem. To maintain good seed stocks, he stressed complete isolation from other varieties to prevent contamination by natural crossing. Peebles (1942) considered 1 mile as a sufficient isolation distance.
Ware (1927) stated that the amount of natural crossing, providing all other factors were the same, would be in direct proportion to the number of insects capable of carrying pollen. He made observations on the cross-pollination of cotton in two areas - at Scott, Ark., where the cotton acreage was large, and at Fayetteville, Ark., where there was little cotton in the vicinity other than the breeding plots. Honey bee colonies were fewer at Scott than at Fayetteville, the number of pollinating insects visiting the cotton flowers was much smaller, and less than 1 percent hybrids were obtained, compared to 40.9 percent hybrids at Fayetteville. Ware (1927) concluded that there was a close association between the presence of honey bees and other pollinating insects and the amount of cross-pollination obtained. Afzal and Khan (1950) reported 2 percent natural crossing with seven or eight visits per flower by insects daily, principally Apis dorsata Fab., Anthophora confusa Smith, and Elis thoracica Lepeletier.
Stephens and Finkner (1953), considering the beneficial effect of insect cross-pollination in the possible production of hybrid cotton, indicated that cross-pollination in different areas ranged from 5 to 50 percent or more, which they associated with differences in the effective bee population. They concluded that even in the area where the higher percentage of crossing occurred, the number of bees did not keep pace with the number of flowers available, so that the flowers were worked less effectively during the peak. Their proposed solution was the provision of a supplementary source of bees during the flowering period.
Simpson (1954a) made a survey of the natural crossing across the Cotton Belt and stated that the cooperators in the experiment were "in general agreement concerning the following factors that influence the amount of natural crossing: (1) cotton pollen is relatively heavy and wind is not an agent in pollen dispersal; (2) therefore, the amount of natural crossing in cotton is determined by the number of insect pollinators present in relation to the number of cotton flowers; and (3) intercrossing may be affected by the flowering habits of the varieties grown, by the abundance of unlike -pollen, by location of the fields in relation to insect habitats, by flowering periods of other plants attractive to insect pollinators, by distance between unlike varieties, by topography and barrier crops, and by other environmental, climatic and biotic factors."
Further in the same paper, he stated that natural crossing had heretofore been considered a handicap in breeding programs and a hazard to be avoided, but that a beneficial intermingling of unlike genotypes could be obtained by increasing the population of insect pollinators. This was supported earlier by Brown (1927). Conversely, Harland (1943) proposed that a completely self-fertile cotton might be grown, a direction that some breeders seem to seek, but which is unlikely to result in the most productive type.
Simpson (1954a) also stated: "Cotton is a partially cross-pollinated plant, thus some degree of heterozygosity is maintained indefinitely when open pollinated seeds from an original F1 population is continued on through F2, and F3, and subsequent generations. The relative proportions of selfing and outcrossing determine the amount of hybrid vigor retained in later generations." He hinted that this relationship could be utilized advantageously in current breeding programs. Kalyanaraman and Santhanam (1957) also hinted that breeding programs such as the mass pedigree method (Harland 1949a,b) should be modified to allow increased genetic plasticity through open pollination and bulk methods of breeding.
Knight and Rose (1954) proposed that one stage of the cotton breeding program be conducted in an area of "high natural crossing" for improvement of the variety. This proposal has not been adopted in the United States, although it and the other statements would indicate that in current cotton breeding systems the breeders should strive for a high bee population in their seed increase fields. Somewhat similar usage of pollinating insects has been suggested by Simpson (1954a) and Turner and Miravalle (1961). The absolute dependence on pollinating insects for success in the Knight and Rose (1954) method was stressed by Bhat (1955), who concluded that natural crossing could result in deterioration of the variety, "not as a result of cross-pollination as is often fallaciously argued but because of its inadequacy." Al-Jibouri (1960) seemed to agree, for after reporting an average of only 0.47 percent crossing in Iraq, he concluded that this inbreeding could result in deterioration of the variety.
It stands to reason that the grower would want to preserve heterosis through favorable gene combinations, or crossing of proper types, and refrain from allowing crossing with inferior or unwanted types.
Many cotton breeders seem to believe that the natural crossing in an area, once established, does not vary from year to year. Thus, they speak of areas of high natural crossing without considering that the factors responsible for crossing may be considerably altered from season to season or even within the season. Humbert and Mogford (1927) stated that cross-fertilization in cotton will vary from 2 to 20 percent and will not average over 15 percent under normal conditions. The "normal" conditions were not described. Fikry (1931) also indicated that natural crossing was a rather fixed 4 percent. Sappenfield (1963) reported a range of 1.0 to 32.2 percent (average, 13.6 percent) natural crossing in Missouri; Simpson (1950)25, a range of 3.3 to 90 percent in his Cotton Belt survey; while Thomson (1966) reported 1 to 2 percent natural crossing in the Ord Valley of northwest Australia. He noted that there were no honey bees in the valley and that insecticides were applied weekly or more often, which, he stated, undoubtedly suppressed any wild bee activity.
Natural crossing, once considered a hazard to be avoided, now is being presented as a tool to be utilized in the development of superior varieties. Also, with the high degree of efficient transportation of apiaries by beekeepers from one area or crop to another, there is no longer assurance that a location will have the same pollinator population from year to year or even throughout the season. This alteration in pollinator population is further accentuated by applications of toxic pesticides to crops, which may damage, destroy, or cause removal of the majority of the pollinators in a given area.
In the "natural pollination" studies made by cotton breeders, none have indicated the relative number of pollinators or visits responsible for the crossing.
___________
25 SIMPSON, D. M. MEMO TO: COOPERATORS IN NATURAL CROSSING TESTS (RESULTS OF 1949 TESTS). Tenn. Agr. Expt. Sta. and U.S. Cotton Field Sta., Knoxville, 3 pp. 1950. [Mimeographed.]MOTES IN COTTON AND THEIR SIGNIFICANCE:
Cotton ovules that fail to develop into seeds with well-developed ginnable fibers are termed "motes." The motes in a cotton crop represent a loss in yield. If 15 to 20 percent or more of the ovules fail to produce seeds with ginnable fibers, the potential yield is lowered just that much (Pearson 1949a). Some structures at the base of certain locks of cotton have the appearance of motes, but they are not derived from ovules. They are termed "false motes." The failure to recognize them as false motes can contribute to an error in estimating the potential seeds that fail to mature (Pearson 1949b).
Rea (1928) reported 6.5 percent motes at the apex of the locks of 'Anton' (upland) cotton and 25.3 at the base, with 11.1 and 38.5 percent, respectively, in 'Bolton' cv. He concluded that this difference might be due to incomplete fertilization. Afzal and Trought (1934) also concluded that mote formation may be due to defective pollination. Rea (1929) found that the higher the percentage of motes, the smaller the boll and the lower the yield of cotton. Hughes (1968) studied motes in 'Bar 14/25', and concluded that most of the motes occurred near the base of the lock in ovary positions 1 and 2. Porter (1936) stated, "To the extent that motes represent unfertilized ovules, it is natural that fewer motes would be found near the apex of the lock, as the ovules near the top of the ovary are favored in fertilization, being reached first by the pollen tubes. If the quantity of pollen deposited on the stigmas is scanty, or much of it defective, there would be a correspondingly small chance of the lower ovules being fertilized." Pearson (1949a) also noted that the number of motes increased with the number of ovules, again an indication of inadequate pollination.
Not only does inadequate pollination contribute to reduced yield through failure of ovules to develop, but mote formation could result in another type of loss. It is easily demonstrated that when a ripe lock of cotton is pulled from the burr, the lock is most likely to break at the location of a mote, and when two motes are adjacent to each other, a break is almost certain. If such a break should occur in the mechanical harvesting of cotton, the remaining portion of the lock would likely be missed by the machine and either lost or only salvaged later at reduced quality during complete destruction of the burr.
The preponderance of bolls with motes in commercial fields indicates the loss of yield being experienced. If the plant "makes up" for motes by setting more bolls, harvest is delayed. The quality of late- developing lint is likely to be inferior to earlier lint.
SHEDDING:
The cotton plant frequently sheds half or more of its fruit. Some of this shedding occurs in the bud stage, but the peak occurs about 5 or 6 days after flowering, then tapers off toward the full-grown boll stage. Many factors contribute to shedding, including humidity, temperature, soil-water conditions, genetic factors, diseases, insects, mechanical injury, and inadequate pollination (Beckett and Hubbard 1932, Brown 1938, Eaton and Ergle 1953, and Lloyd 1920). Rainfall during the day the flower is open can damage the pollen and cause shedding although the amount of shedding from this factor is minor. Kearney (1923) doubted that deficient pollination and fertilization were the primary reasons for boll-shedding in Pima cotton at Sacaton, Ariz. However, Kaziev (1964) showed that bee pollination caused a decrease in the shedding of five common cultivars of upland cotton. McGregor et al. (1955) also reported reduced shedding of 'Pima S-1' cotton visited by bees as compared to plants caged to exclude bees. Occasionally, during periods of extremely hot weather, the cotton flower will fail to produce or dehisce viable pollen. When this occurs, the flower sheds unless pollen is brought to it from another flower by insects.
Many factors, therefore, contribute to shedding by cotton, one of which is inadequate pollination.
NEED FOR POLLEN TRANSFER WITHIN OR BETWEEN PLANTS:
When the cotton pollen grains contact the stigma, they germinate tubes that grow down the "conducting tissue" in the center of the style (Arutinnova 1940). When a tube reaches the ovary, the sperm enters and fertilizes an ovule. About 50 ovules must be fertilized if a full complement of seeds is produced; therefore, at least 50 viable pollen grains must contact the stigma. The stigma is normally receptive to the pollen by the time the flower opens or before (Loden et al. 1950), but receptivity drops sharply after about noon (Janki et al. 1968). Because of this limited time span, most hand pollination in cotton breeding work is made during the forenoon.
The majority of the flowers on a cotton plant are largely self- fertile and, to varying degrees, self-pollinating (Ewing 1918, Kearney and Harrison 1932). The method, time, and type of pollination of the stigma influence the degree of self-pollination. Flowers that receive pollen on the whole surface of the stigma yield more seeds per boll than those pollinated at the base of the stigma (Kearney 1926). The base of the stigma affords less favorable condition for pollen germination or growth than the apex (Iyengar 1938, Kearney 1923). Pollen from anthers on the lower part of the stamina column is best (Trushkin 1956). Repeated applications of pollen to the stigma, such as repeated bee visits, are also helpful (Finkner 1954). Therefore, for best pollination of the stigma, repeated applications with an abundance of pollen from the basal anthers on the tip of the stigma insures the highest percentage of fruit set and maximum cotton production.
The pollen-laden anthers that touch the stigma, resulting in self- fertilization, usually contact only the base of the stigma. When this occurs, self-pollination is assured, but maximum fertilization within the ovary of that boll is not usually accomplished. Arutiunova and Gubanov (1950) indicated that "pollen seems to stimulate pollen" so that increased amounts of it on the stigma increased the percentage of germinating pollen tubes, further insuring ovule fertilization.
Rose and Hughes (1955) increased yield of 'Bar 7/8.2' by 11 percent over naturally pollinated flowers by brush pollination of stigmas. The increase resulted from more bolls set. Kohel (1968) also obtained more seeds per lock and per boll from unbagged open flowers, or hand-pollinated open flowers than from bagged or emasculated flowers. Guseinov and Muktarov (1963) showed that cross-pollination within the variety resulted in increased production.
Some pollens are too weak to compete with self-pollen. For example, when McGregor et al. (1955) grew equal numbers of 'A-44' and 'Red Acala' plants in cages with honey bees, only 2.31 percent of the offspring of 'A- 44' were hybrids, whereas 44.0 percent of the 'Red Acala' offspring were hybrids.
As the pollen tube grows down the style, its nucleus moves a few microns ahead of the sperm (Jensen and Fisher 1968). The sperm and contents are discharged into the germ sac of the ovule in 16 to 32 hours (Gore 1932, Kearney 1923, Saakyan 1962). Surplus pollen tubes that penetrate the ovary are eventually assimilated without damage (Linskens 1964), so there is no damage from surplus pollen on the stigma.
According to Arutiunova (1940), the pollen tube begins to form more quickly if the pollen grain is from a genetically different cultivar. For example, tubes in cross-pollinated flowers were visible within 5 to 10 minutes after the pollen was placed on the stigma, but tubes from self pollen did not appear until after 60 to 150 minutes. Also, tubes from pollen placed on the tip of the stigma grew faster than those from pollen placed on the base of the stigma.
Arutinnova (1940) also studied the effect of the number of pollen grains and the kinds of intravarietal pollen on pollen tube growth in both upland and G. barbadense cottons and reported that in the G. barbadense cottons twice as many tubes reach the ovary with cross-fertilization as with selfed flowers. Except for one cultivar, pollen of upland cottons grew better on its own stigma than on others. In many of the selfed flowers, no pollen tubes developed, but in crossed flowers this failure was rare.
This means that more self pollen must be deposited on the stigma than mixed or cross pollen. The sooner the tube reaches the ovary and fertilizes an ovule, the less the likelihood that the fruit will shed. Arutiunova and Gubanov (1950) also concluded that an increased number of pollen grains on the stigma increased the percentage of germinating grains and tube development. Arutiunova and Kanas (1955) concluded that cross-pollination within the cultivar insures the best seed development. In other words, a well cross-fertilized stigma tip is most likely to result in the best fertilization of the ovules in the ovary.
There seems to be constant warfare within the plant between setting and shedding of fruit. Anything that can be done to influence the battle in favor of fruit setting tends to increase production. The earliest possible thorough application of pollen on the tip of the stigma to insure speediest arrival of the tubes in abundance within the ovary would insure maximum set.
BENEFITS DERIVED FROM INSECT POLLINATION OF COTTON:
The benefits derived by cotton from insect pollination have been cited by numerous workers, largely in Russia, where much attention has been given to this subject, but also in Egypt, India, and the United States. Meade (1918) was the first to call attention to the fact that cotton at San Antonio, Tex., should benefit from insect pollination. He applied supplementary pollen by hand to open flowers that may or may not have been visited by pollinating insects and increased the set of 'Durango' cotton flowers by 10.96 percent and 'Acala' flowers by 5.31 percent. His results convinced him that pollination of cotton by honey bees should increase production.
Kearney (1921) was stimulated by Meade's work to compare naturally pollinated Pima flowers with flowers that received supplementary pollination by hand. At Sacaton, Ariz., where the cotton was relatively isolated and insect pollinators were prevalent, he obtained no significant increase in set of flowers or seed produced. However, at Phoenix, in a large cotton-growing area, he obtained only 1,157 seeds per 100 naturally pollinated flowers, but 1,526 seeds per 100 flowers that received supplementary pollination. As a result, he, too, recommended the keeping of bees in cotton fields for their pollination service. Later, Kearney (1923) concluded that Pima cotton production in the Salt River Valley of Arizona could be substantially increased (about 32 percent) if honey bees were kept around the cotton fields.
The rather comprehensive and convincing research by Shishikin (1946, 1952) apparently awakened his countrymen to the value of bees to cotton. His work was followed by that of numerous workers but especially certain leaders in this work: Kaziev (1955, 1956a, b, 1958, 1959a, b, 1960, 1961a, b, 1963,1964, 1967), Kuliev (1958), Minkov (1953a, b), Radoev (1963, 1965), Radoev and Bozhinov (1961), Skrebtsov (1964), and Trushkin (1956,1960a, b).
In Peru, Mercado Mesa (1956) concluded that insect pollination was of no value to 'Tanguis' cotton (G. barbadense). In a more thorough test Dulanto Bartra (1958) showed that 51.7 percent of the flowers shed if they were not visited by the bee Melitoma euglossoides Lepeletier and Serville compared to only 32.2 percent of flowers visited by this bee.
Shishikin (1946) was the first to use the term "saturation pollination" - the uniform distribution of colonies of honey bees among cotton fields. He showed that saturation pollination, at the rate of one- half colony per acre, increased production of cotton 19.5 percent more than areas dependent upon only local pollinators. The increase over cotton grown in cages, excluding all insects, was 43 percent. In his more complete report, Shishikin (1952) stated that 4,130 to 5,000 colonies were used, at the rate of one colony per hectare, and distributed in groups 1 km apart. The tests dealt with "the old variety, '114', and the new variety '1298'." During the tests, the natural crossing caused by the honey bees varied from 26 to 43 percent. He concluded that the wild pollinators were "far from being able to assure cross-pollination of cotton plants."
In a sense, Babadzhanov (1953) duplicated the test by Kearney (1923) and obtained a similar benefit from supplemental pollination. He reported that it increased the boll set of cultivar '108-F' by 30 percent, the raw cotton per boll by 5 to 10 percent, the seed germination from 93 percent in selfed seed to 98 percent in cross-pollinated seed, and decreased the motes by 12.5 percent. Ter-Avanesyan (1952) showed that crossing within the cultivar varied with the cultivar tested: 8.4 percent in 'Sreder', 14.4 in '8582', and 22 percent in '915'.
These tests were supported in theory by Miravalle (1964), who compared the effects of bulked pollen from several plants with pollen from one flower of the same selection. He found that 76 percent of the bolls set, with 34.47 viable seeds per boll, when the flowers were pollinated with bulked pollen, but only 70 percent set, with only 27.07 seeds per boll, when the flowers were pollinated with pollen from one flower of the same plant.
McGregor et al. (1955) studied the effect of bee pollination upon upland and Pima cotton in cages, some of which contained a colony of honey bees (figs. 92, 93). In their test, 'Pima S-1' produced 24.5 percent more cotton in cages with bees than in cages without bees. This increase was caused by the set of more bolls, with more seeds per boll. The presence of bees did not increase total production of upland 'A-33' or 'A- 44', but the crop set earlier. In an area with a short season, this effect would doubtless be reflected in a greater total yield. Also, the cotton was handpicked, with extreme care taken to collect every seed. If machine harvesting had been used, doubtless more cotton would have been collected where there were fewest motes - in the bee cages. The lock usually breaks if motes are prevalent, and the remaining lint remains unharvested in the base of the burr.
In this test and others, McGregor and Todd (1955, 1956) noted that a decided difference developed before the end of day in the appearance of the petals of the upland cotton flowers visited by bees as compared to those not visited. By mid-afternoon, the visited ones began to change color and wilt, and the petals formed a tight cylindrical roll; whereas, in those flowers not visited by bees, the petals stayed white and open until sunset then became limp and adhered to each other like pieces of wet paper (fig. 94).
Mahadevan and Chandy (1959), using cultivars 'M.U.I.' and 'M.C.U.2' in India, obtained 23 to 34 percent and 40 to 53 percent, respectively, more cotton in open plots than in plots caged to exclude bees. They did not have plots caged with pollinating insects. This leaves unexplained the possible effect of the caging on the plant. Sidhu and Singh (1962), also in India, compared production in cages with Apis indica [cerana] and A. florea and in cages without pollinators and obtained an increase of 17.45 to 18.98 percent in favor of the pollinating insects. The increase was attributed to more and larger bolls.
In the United Arab Republic, Wafa and Ibrahim (1960) also obtained 22.4 percent more 'Ashmouni' cotton with honey bee pollination.
Skrebtsov (1964) obtained 33 percent increase in raw cotton by cross-pollination within the variety with honey bees, and showed that the bees improved hybrid vigor.
Radoev and Bozhinov (1961) obtained 10.6 to 24.4 percent greater yield from flowers freely visited by bees than from flowers tied to exclude insects during flowering. There were 0.5 more seeds per boll, fewer motes, and better seed germination. Radoev (1963) stated that freely pollinated plants set 11.04 percent more cotton than isolated plants, with more sound seed and better germination. Radoev (1965) concluded that the honey bee is the most important insect in Russia in the pollination of cotton, even though only 18 percent of the floral visits contribute to pollination.
Minkov (1953b) studied the effect of pollination on Russian cultivars '611-b' and '108-F', which were visited primarily by wild bees. He found that exclusion of pollinators increased the number of motes.
These tests indicate that the value of insect pollination is not limited to any particular area, species, or cultivar.
The material in the reports by the various authors previously mentioned as well as numerous others, leaves little doubt that cotton is benefited by bees in terms of greater lint and seed production, earliness of harvest, fewer motes, better lint, better germination, and improved qualities in the offspring.
Trushkin (1960a) concluded that the use of bees on cotton must be considered not only possible but expedient. Trushkin (1960b) stated, "The time has come to fully exploit [utilize] honey bees for purposes of obtaining high cotton yields and improving seed quality ...."
[gfx]
FIGURE 92. - Pollination studies involving honey bees on caged cotton plants.
FIGURE 93. - Author taking data for pollination studies on tagged Pima cotton flowers.
FIGURE 94. - Effect of insect pollination on cotton flowers. Photograph taken 4:30 p.m. of the day these 2 flowers opened. Flower on left was in a bee cage. Its petals had changed from cream to pink and tightened into a tubular shape. Flower on right was in a no-bee cage. Its petals remained cream-colored and flared until sunset.HYBRID VIGOR IN COTTON:
The accentuated effect of cross-fertilization is referred to as heterosis or hybrid vigor. It can result from interspecific (between species), intraspecific (within species), or intervarietal crossing. The possible utilization of hybrid vigor in cotton has been of considerable interest to cotton breeders since Mell (1894) showed that some cotton hybrids exceeded their parents in certain characteristics. Although Kottur (1928) concluded that selfing of G. herbaceum plants for 12 generations had no injurious effect, and Harland (1943) proposed the breeding of a cotton immune from natural crossing, Brown (1942) and O'Kelly (1942) independently concluded that inbreeding of upland cotton reduced production and caused fewer flowers and smaller bolls. Simpson and Duncan (1953) also showed that cultivars selfed for 10 years produced 15 percent less cotton than the original plants. As a result, breeders now generally agree that too much inbreeding is detrimental. Instead, they strive for or desire some degree of outcrossing but, so far, have not been able to control it. The subject of hybrid vigor in cotton was thoroughly reviewed by Loden and Richmond (1951).
Hybrid vigor in cotton has been observed in interspecific crosses as well as in crosses between varieties within the species. Fryxell et al. (1958), Hutchinson et al. (1938), Marani (1967), Stroman (1961), and Ware (1931) in particular showed that crosses between G. barbadense and G. hirsutum were much more productive than either parent. Because of the differences in the characteristics of the lint of the two species, it frequently has objectionable qualities in the hybrid. This problem is less likely to arise in intraspecific hybrids where considerable hybrid vigor has also been shown.
Kime and Tilley (1947) made numerous crosses of commercial cultivars of upland cotton and showed an increase in production of the F1 ranging from 7 to 20 percent. They doubted that hand cross-pollination to produce the hybrid seed was practical but considered production feasible in areas where "a high percentage of crossing normally occurs." Kohel and Richmond (1969) showed that significant heterosis could be obtained in areas of high natural crossing. Patel and Patel (1952) indicated that in India hand-pollination might be practical. Thakar and Sheth (1955) proposed that the government subsidize hybrid cotton seed production. Simpson (1948) obtained increases in yield from crosses of upland cultivars ranging from 5.7 to 44.2 percent. He, like Meade (1918) and Kearney (1923), also recommended that honey bee colonies be placed around the fields, knowing that hybrid seed could only be produced in quantity with pollinating insects (natural crossing) (Simpson 1954b).
Hybrid vigor has been shown within upland cotton by numerous workers (Barnes and Staten 1961, Christidis 1955, Galal et al. 1966, Hawkins et al. 1962, Lee et al. 1967, and Turner 1953a, b ).
Miller and Lee (1964) reported larger bolls with higher lint yields. Muramoto (1958) showed increases in yield, lint percentage, lint index, and seeds per boll. Ter-Avanesyan and Lalaev (1954) reported yield increase, bolls ripening 5 to 6 days earlier, and some resistance to Verticillium wilt. Trushkin and Truskina (1964) also reported that supplementary pollination by bees increased resistance to wilt, fungal root rot, and Xanthomonas maluacearum. Kaziev (1961b) reported earlier germination of hybrid seed and wilt resistance.
Wanjura et al. (1969) showed the importance of early emergence of the seed to plant survival and yield. For plants emerging on the fifth, eight, and 12th days, the survival was 87, 70, and 30 percent, and the relative yield of the plants was 100, 46, and 29 percent respectively.
The value of the hybrids is not in the mere mixing of plant types in the field, such as might occur in certain breeding programs without insect pollination. Richmond and Lewis (1951) showed that from the standpoint of yield nothing was gained by growing a mixture of seed types in a pure stand although such a mixture, they indicated, might supply a mixture of fibers not obtainable from a lone commercial cultivar.
A method tried by Peebles (1956) consisted of planting alternate rows of 'Pima 32' and 'Pima S-1' and the use of saturation pollination by honey bees. He showed the economic feasibility of this method, but it was not accepted by the industry. An attempt was made by a commercial company (DeKalb Agricultural Association, Inc. 1961) to produce hybrid cotton, but their supply of pollinating insects was apparently inadequate. Instead of attempting to increase the local supply, they concluded that hybrid cotton seed must be produced "in marginal cotton growing areas where bee activity is great." Turner (1959) proposed the planting of appropriate male-sterile and normal-functioning flower seed mixtures, then reaping the benefit of the hybrid vigor caused by thorough cross- pollination between the plants by a high population of honey bees.
One method frequently proposed (Christidis and Harrison 1955) for utilizing hybrid vigor involved male-sterile plants. Allison and Fisher (1964), Fisher (1961), Justus and Leinweber (1960), Justus et al. (1963), Meyer and Meyer (1965), Turner (1948), and Weaver and Ashley (1971) have reported the presence of male sterility in cotton. At one time, the creation of male-sterile plants with a chemical "gametocide" (Sodium 2, 3-dichloroisobutyrate) looked promising (Eaton 1957, McGregor 1958, Meyer et al. 1958, and Rohm and Haas 1958); however, subsequent testing failed to establish its reliability. It also created some female sterility; therefore, its use was discontinued.
Stith proposed (1970) the use of cytoplasmic male-sterile stocks and restorer genes in cotton cross-pollinated by bees. Kohel and Richmond (1962) showed that bees should function satisfactorily in the production of cotton on male-sterile plants. Meyer (1969), commenting on the progress made with cytoplasmic male sterility, stated that the basic plant work has been done, but the bee breeders and their bees still have a lot of work to do. She concluded that the most critical problem in the production of hybrid cotton appeared to be in finding some way to get the male-sterile flowers pollinated.
Hybrid vigor in cotton offers possibilities for increasing cotton production to a new plateau, if insects can be used as the cross- pollinators. Tests mentioned herein show this is possible. The problem is to find the proper cotton combiners and the best utilization of pollinating insects.
Pollinators:
There is agreement that cotton is not wind pollinated (Balls 1915), that all pollen transport outside of the flower requires an active vector, and that "bees" are the best pollinators of cotton. Tsyganov (1953) stated that "bees on the cotton flower are not guests but thoroughly adapted symbionts because they feed and rear their young on the products gathered from the flowers."
The bees most frequently mentioned are the bumble bees (Bombus spp.), honey bees (Apis dorsata, A. florea, A. indica [cerana], and, most frequently, A. mellifera), and the solitary groundnesting Melissodes spp. Other hymenoptera sometimes mentioned include Anthophora spp., Elis thoracica Lepeletier, Halictus spp., Megachile spp., Melitoma euglossoides Lepeletier and Serville, and Nomia spp. Numerous species from several other orders of insects sometimes find their way into cotton flowers, but as Simpson and Duncan (1956) stated, pollen distribution is essentially a "put and take" procedure, and unless the insect consistently visits large numbers of cotton flowers it is relatively ineffective as a pollinator.
In the United States, the bumble bee, honey bee, and Melissodes bees are considered most important as pollinators of cotton (Allard 1910, 1911a, b, Butler et al. 1960, Kearney 1923, McGregor 1959, McMillian 1959, Stephens and Finkner 1953, Theis 1953). In Russia, the honey bee is undisputedly considered the most important. In India, Apis spp., Elis thoracica, and Anthophora spp. have been mentioned (Sidhu and Singh 1961, Khan and Afzal 1950). In Egypt, the honey bee is most commonly seen (Wafa and Ibrahim 1957, 1959).
BUMBLE BEES:
Brown (1927) stated: "Large lubberly bumble bees that get pollen all over their bodies and rub against the stigma of every flower they meet are doubtless the best." With this there is no disagreement. Many of the cotton researchers concerned about insect visitation to cotton flowers in the Cotton Belt east of the Brazos River consider the bumble bee most important (Allard 1910, 1911a, Loden and Richmond 1951, Stephens and Finkner 1953, Theis 1953).
The visits to the plant by bumble bees are predominantly within the flower. Because of its size, the bumble bee can scarcely enter the flower without depositing pollen on the stigma and picking up more from the anthers. Because the nest is provisioned with both nectar and pollen, the bumble bee makes numerous collecting trips to the flowers. Also, bumble bees are colonial and under favorable conditions the population within the nest may increase so that numerous individuals from one nest will be foraging simultaneously in a field.
In isolated cotton test plots in North Carolina, it is not unusual to find a bumble bee on every plant and at times in every flower. Under such conditions, their effectiveness as pollinators could not be surpassed. Incidentally, Dulanto Bartra (1958) reported up to three or four Melitoma euglossoides in a single flower near the nesting sites, but they were very scarce farther away. By contrast, in the western half of the Cotton Belt, an entire day might be spent in a large cottonfield without seeing a bumble bee. Here, they are of no importance whatever as pollinators of cotton.
Bumble bees are colonial only through the active season. The nest is abandoned in the fall, the males die, and the females go into hibernation. Each female that survives hibernation establishes a new nest in the spring. Nests are only established if suitable nesting sites can be found. The colony in the nest then faces numerous hazards throughout the season, such as lack of a continuous source of fresh nectar and pollen, diseases, pests, pesticides, and other environmental factors or agricultural practices. For these reasons, bumble bees are not always present in adequate numbers when desired, and their numbers cannot be increased as desired. The culture of bumble bees for the pollination of cotton holds little promise.
MELISSODES BEES:
The Melissodes bee frequently constructs its nest in the soil in the cottonfield. It visits cotton flowers as a preferred host plant and rarely if ever visits extrafloral nectaries. It provisions each cell of its nest with a l/5-inch pellet of pollen and nectar. The female spends the night in her subterranean nest. She rapidly visits blossom after blossom of either upland or Pima cotton and will cross over from the one to the other on a single trip. Under natural conditions, a single bee may make as many as 200 floral visits in a day (Butler et al. 1960).
Melissodes bees are quite seasonal, therefore, they may be plentiful during one part of the flowering season but rare later in the same season. They are adversely affected by pesticides applied during the daytime, but the females may escape damage from nighttime applications. Little is known about the adverse effects on them of insecticides, soil cultivation, irrigation, or crop rotation. No way is known to increase these bees when desired.
HONEY BEES:
In contrast to bumble bees and Melissodes bees, honey bees show a preference for the extra-floral nectaries of cotton and often seem reluctant to enter the cotton flower. When a honey bee enters a cotton flower, it may emerge coated with pollen, then alight on a leaf, and comb much of the pollen off without attempting to pack it in the pollen baskets on the hind legs. However, all of this pollen is not removed, and a familiar sight, where bees are working cotton, is their incoming at the hive entrance coated with cotton pollen. Radoev (1965), in Russia, stated: "The honey bee is the most important insect in the pollination of cotton." As shown in other places herein, its value in the United States would appear to be in proportion to its use and concentration on the cotton.
At times, honey bees collect small amounts of cotton pollen and transport it to the hive (fig. 95). This usually occurs only when no other pollen is available for the bees. Minkov (1956) concluded that honey bees can collect cotton pollen but seldom do so. On the other hand, Kaziev (1956a, 1964) stated that 15 to 25 percent of the bees were collecting pollen when the average colony was storing 2 to 5 pounds of honey per day. The bees were collecting the pollen from 8 a.m. to 4 p.m. Whether he referred to their actually collecting the pollen in their pollen baskets or whether they were merely entering the hive with pollen on their bodies, indicating that they had been inside the cotton flowers, is not clear.
Pollen collection is not always dictated by supply and demand. Honey bee colonies have been observed in Arizona by the author (unpublished data) and Grout (1955) showing every evidence of pollen deficiency, although these colonies were surrounded by hundreds of acres of both species of cotton in flower. Later in the season, the bees in the same location collected cotton pollen freely. No reason could be determined for this strange behavior. At all times, the honey bees which were concentrated at the rate of one colony for each acre of cotton, were collecting both floral and extrafloral nectar.
In 1957 (unpublished data), I counted the honey bees in cotton flowers on each of five farms at Shafter, Calif., and four farms near Mettler Station, about 50 miles to the south. In both areas, the only cotton grown was 'Acala 4-42', and in both areas cultivation was large scale and dependent on irrigation water. Near Shafter, water was plentiful, and much of the land was devoted to cotton and alfalfa hay production. There were few apiaries near Shafter. Near Mettler Station, the water supply was acute; therefore, the growers devoted some of their land to alfalfa seed production, which required less water than cotton. The alfalfa seed fields and cotton fields were interspersed. About 50,000 colonies of honey bees had been transported into the Mettler area to pollinate about 20,000 acres of alfalfa seed. In the Shafter cotton fields, only 13 honey bees were observed in 1,000 cotton flowers (1.3 bees per 100 flowers), but at Mettler Station 158 were counted (15.8 bees per 100 flowers). One unidentified wild bee was seen.
All cotton fields within one-quarter mile of the alfalfa fields had 20 or more honey bees per 100 flowers. This proved that honey bee populations can be built up to provide "saturation pollination" or the "10 bees per 100 flowers" shown by McGregor (1959) to be sufficient to provide thorough coverage of the stigma with pollen.
In an 80-acre 'Pima S-1' cottonfield at mid-season in Arizona, Johansson (1959) dusted fluorescent pigmented particles in a single, newly opened flower. The following day he recorded the percentage of day-old (closed) flowers showing the presence of such particles (brought the previous day) in relation to distance from the treated flower. There were 212 colonies of honey bees within or along the borders of this field (fig. 96), but few other pollinating insects were active where the test was conducted. His results were: 0 to 50 feet (40.5 percent), 50 to 100 feet (14.0 percent), 100 to 150 feet (3.5 percent), and 150 to 200 feet (1.6 percent). This showed that when honey bees are present in sufficient numbers, they can effectively distribute such particles (and pollen grains) from flower to flower.
Kohel and Richmond (1962) concluded that a single insect visit to a cotton flower is not enough for complete pollination. Ter Avanesyan (1959) showed that 600 to 1,000 pollen grains on the stigma is the minimal "norm" quantity of pollen required. Shoemaker (1911) and Minkov (1953b) mentioned the commonly observed characteristic of the honey bee in alighting on the corolla rim, after which it crawls down the petal with its back to the anthers, rarely touching the stigma. However, when there is sufficient honey bee traffic into the cotton flower some of the individuals "get careless" and alight upon or crawl over the stigma, giving it a liberal coating of pollen in the process. The secret of successful pollination of cotton with honey bees seems to be in having sufficient visitation so that the bees are "forced" to visit the flowers, and considerable bee traffic into and out of the flower results.
Pollinating bees are an obstacle to most cotton breeders attempting to develop pure lines. Each cotton blossom from which the breeder desires seed must be enclosed or isolated in some way, otherwise the pollinating insect may dilute the line by bringing pollen to it from another type of cotton plant. This is particularly true if the breeder is using the individual plant selection method where he breeds offspring from a single plant. However, Brown (1942) and Simpson and Duncan (1953) have shown that continual inbreeding causes a decrease in productiveness not fully compensated by gains in other properties. As a result, Harland (1949a) proposed his mass pedigree selection system in which outcrossing within the mass is permitted, and Knight and Rose (1954) proposed a modification in which there is the initial selfing generation, then a selection of progeny rows, the seeds from which are bulked and grown in an isolated area with the highest pollinator population that can be obtained.
Thus, the pollinating insect changes from a "harmful insect in the development of the plant selection to "beneficial" insect in the later stages of the program but there still remains within the minds of many cotton specialists an aura of animosity toward these insects. This has been coupled with the fact that bees are not "necessary" in the production of cotton. The evidence strongly indicates that for the best interest of the grower the pollinating insects should be protected and their presence encouraged in breeding and seed increase programs and in the production of bulk cotton.
Although the use of honey bees for systematic or saturation pollination of cottonfields is practiced to considerable extent in Russia, resulting in increase and/or improved productivity, the American grower has tended to strive for his increased production through the use of pesticides and other agronomic practices and has given little heed to the beneficial insects. Some growers, after observing that areas near apiaries are frequently more productive, feel that bees are of some value. Others, fearing that their pesticide program might damage the bees and result in legal action by the beekeeper, discourage the keeping of bees near the cottonfields. The request for or the rental of bees to pollinate cotton in the United States is extremely rare.
The cotton plant may flower for 2 months or more; however, Buie (1928) showed that the majority of the flowers that set fruit appeared within 3 to 4 weeks.: the bees were concentrated on the cotton for this period with the use of harmful pesticides curtailed, the bees could perform their pollination service and escape pesticide damage.
[gfx]
FIGURE 95. - Honey bee collecting cotton pollen
FIGURE 96.- Honey bee colonies beside cotton field.Pollination Recommendations and Practices:
Meade (1918), Kearney (1923), and Stephens and Finkner (1953) recommended the keeping of honey bees near cottonfields, but no ratio of bees per flower or colonies per area was indicated. Shishikin (1952), who studied the effect of bees on about 5,000 acres of cotton in Russia, recommended one colony per acre with the colonies grouped about 0.6 mile apart. McGregor and Todd (1955) suggested one colony per acre. Avetisyan (1965, ch. 5, pp. 209-248) suggested 0.5 to 1.0 colony per hectare (one colony per 2.5 to 5. acres). Glushkov and Skrebtsov (1960) stated that with 4.9 and 6.6 colonies per hectare (2.0 to 2.5 colonies per acre) the cotton production was increased 20.9 and 45 percent, respectively, over the control areas.
The colonies-per-acre ratio (from one-fifth of colony to five colonies per acre have been suggested) is doubtless influenced by the acreage involved, competing crops, and colony strength. The ratio of 10 bees per 100 flowers suggested by McGregor (1959) is a more realist ratio than colonies per acre. Quite probably, a low population of honey bees contributes little or nothing to pollination. A method of maintaining a high population should be considered when honey bees are used. Although not an ideal pollinator of cotton, the honey bee is the only pollinator that can be manipulated on cotton.
LITERATURE CITED:
ANONYMOUS.
1972. COTTON - WORLD STATISTICS. Internatl. Cotton Advisory Com. Quart. Bul. 25(9, part 2): 1 - 144, Washington, D.C.AFZAL, M., and KHAN, H.
1950. NATURAL CROSSING IN COTTON IN WESTERN PUNJAB. Agron. Jour. 42: 14 - 19, 89 - 93, 202 - 205, 236-238.______and TROUGHT, T.
1934. MOTES IN COTTON. I. PUNJAB-AMERICAN COTTON. Indian Jour. Agr. Sci. 4: 554-575.AL-JIBOURI, H. A.
1960. NATURAL CROSSING IN COTTON IN IRAQ. Agron. Jour. 52: 53.ALLARD, H. A.
1910. PRELIMINARY OBSERVATIONS CONCERNING NATURAL CROSSING IN COTTON. Amer. Breeders' Mag.(4): 247 - 261.______ 1911a. SOME EXPERIMENTAL OBSERVATIONS CONCERNING THE BEHAVIOR OF VARIOUS BEES IN THEIR VISITS TO COTTON BLOSSOMS. Amer. Nat. 45(538): Part I, 607 - 622; Part II, 668 - 685.
______ 1911b. PRELIMINARY OBSERVATIONS CONCERNING NATURAL CROSSING IN COTTON. Amer. Breeders' Assoc. Mag. 6: 156 - 170.
ALLISON, D. C., and FISHER, W. D.
1964. A DOMINANT GENE FOR MALE-STERILITY IN UPLAND COTTON. Crop Sci. 4: 548. ARUTIUNOVA, L. G. 1940. [GERMINATION AND DEVELOPMENT OF COTTON POLLEN IN INTRAVARIETAL CROSS-POLLINATION.] Jarovizacija 1(28): 18-22. [In Russian.]______and GUBANOV, G. IA.
1950. [IN REGARD TO THE BIOLOGY OF COTTON FERTILIZATION.] Agrobiologiya 6: 94-99. [ln Russian. ] Biol. Abs. 25(4): 31994, 1951.______and KANAS, M.
1955. [FACTORS WHICH LESSEN THE FORMATION OF MOTES IN COTTON.] Khlopkovodstvo 5(8): 43 - 47. [In Russian.]AVETISYAN G. A.
1965. [BEE POLLINATION OF AGRICULTURAL CROPS.] His Pchelovodstvo, Moskva, "Kolos," pp. 209-248. [In Russian.]BABADZHANOV, F.
1953. [ROLE OF INTRAVARIETAL POLLINATION IN INCREASING THE PRODUCTIVITY OF COTTON.] Khlopkovodstvo 3(7): 34 - 39. [In Russian.]BALLS, W. L.
1912. THE COTTON PLANT IN EGYPT. 202 pp. Macmillan and Co., Ltd., London.______ 1915. THE DEVELOPMENT AND PROPERTIES OF RAW COTTON. 221 pp. Black Ltd., London.
BARNES, C. E., and STATEN, G.
1961. THE COMBINING ABILITY OF SOME VARIETIES AND STRAINS OF GOSSYPIUM HIRSUTUM. N. Mex. Agr. Expt. Sta. Bul. 457, 33 pp.BECKETT, R. E., and HUBBARD, J. W.
1932. THE SHEDDING OF 4-LOCK AND 5-LOCK BOLLS IN UPLAND COTTON. U.S. Dept. Agr. Tech. Bul. 277, 16 pp.BEDFORD, H. W.
1921. THE ASAL OF COTTON AND ITS CAUSES IN THE SUDAN. Gordon Mem. Col. Khartum, Wellcome Res. Lab. Ent. Sect. Bul. 17, 8 pp.BENTON, R.
1937. IN THE LAND OF COTTON. Bees and Honey 18: 364 - 365, 378 - 379.BHAT, N. R.
1955. SOME POINTS DESERVING CONSIDERATION IN THE BREEDING AND PURE SEED MULTIPLICATION OF COTTONS IN INDIA. Indian Cotton Crowing Rev. 9(4): 189 - 196.BROWN, H. B.
1927. VICINISM OR NATURAL CROSSING IN COTTON. Miss. Agr. Expt. Sta. Tech. Bul. 13, 14 pp.______ 1938. COTTON. 582 pp. McGraw-Hill Book Co., Inc., New York.
______ 1942. RESULTS FROM INBREEDING UPLAND COTTON FOR A 10-YEAR PERIOD Amer. Soc. Agron. Jour. 34: 1084-1089.
BUIE, T. S.
1928. THE FRUITING HABITS OF THE COTTON PLANT. Amer. Soc. Agron. Jour. 20: 193 - 201.BUTLER, G. D., JR., LOPER, G. M., McGREGOR, S. E., and others.
1972. AMOUNTS AND KINDS OF SUGARS IN THE NECTARS OF COTTON (GOSSYPIUM SPP.) AND THE TIME OF THEIR SECRETION. Agron. Jour. 64: 364 - 368.______TODD, F. E., McGREGOR, S. E., and WERNER, F. G.
1960. MELISSODES BEES IN ARIZONA COTTON FIELDS. Ariz. Agr. Expt. Sta. Tech Bul. 139, 11 pp.CHRISTIDIS, B. G.
1955. HYBRID VIGOR EFFECTS WITH COTTON. Jour. Genet. 53(2): 225-231.______and HARRISON, G. J.
1955. COTTON GROWING PROBLEMS. 633 pp. McGraw-Hill Book Co., Inc., New York and London.COOK, O. F.
1921. IMPROVEMENTS IN COTTON PRODUCTION. U.S. Dept. Agr. Cir. 200, 12 pp.______ 1932. COTTON IMPROVEMENT THROUGH TYPE SELECTION. U.S. Dept. Agr. Tech. Bul. 302, 62 pp.
DEKALB AGRICULTURAL ASSOCIATION, INC.
1961. DEKALB COTTON SEED. THE COTTON WITH THE BEE IN ITS BREEDING. DeKalb Agr. Assoe., Inc., Athens, Ga., 11 pp.DULANTO BARTRA A.
1958. [THE IMPORTANCE OF THE "BUMBLE BEE", MELITOMA EUGLOSSOIDES LEP. & SERV. IN THE POLLINATION OF THE FLOWERS OF THE TANGUIS COTTON PLANT.] Rev. Peruana de Ent. Agr. 1(1): 6-11. [In Spanish.]DUNLAP, A. A.
1945. FRUITING AND SHEDDING OF COTTON IN RELATION TO LIGHT AND OTHER LIMITING FACTORS. Tex. Agr. Expt. Sta. Bul. 677, 104 pp.EATON, F. M.
1957. SELECTIVE GAMETOCIDE OPENS WAY TO HYBRID COTTON. Science 126: 1174 - 1175.______and ERGLE, D. R.
1953. RELATIONSHIP OF SEASONAL TRENDS IN CARBOHYDRATE AND NITROGEN LEVELS AND EFFECTS OF GIRDLING AND SPRAYING WITH SUCROSE AND UREA TO THE NUTRITIONAL INTERPRETATION OF BOLL SHEDDING IN COTTON. Plant Physiol. 28(3): 503-520.EWING, E. C.
1918. A STUDY OF CERTAIN ENVIRONMENTAL FACTORS AND VARIETAL DIFFERENCES INFLUENCING THE FRUITING OF COTTON. Miss. Agr. Expt. Sta. Tech. Bul. 8, 93 pp.FIKRY, M. A.
1931. NATURAL CROSSING IN COTTON. Roy. Agr. Soc. Egypt, Tech. Sect. Bul. 18, 23 pp.FINKNER, M. D.
1954. THE EFFECT OF DUAL POLLINATION IN UPLAND COTTON STOCKS DIFFERING IN GENOTYPE. Agron. Jour. 46(3): 124 - 128.FISHER, W. D.
1961. MALE-STERILE COTTON FOUND. Prog. Agr. in Ariz. 13(2): 11.FRYXELL P. A.
1957. MODE OF REPRODUCTION OF HIGHER PLANTS. Bot. Rev. 23: 135 - 233.______STATEN, G., and PORTER, J. H.
1958. PERFORMANCE OF SOME WIDE CROSSES IN GOSSYPIUM. N. Mex. Agr. Expt. Sta. Bul. 419, 15 pp.GALAL, H. E., MILLER, P. A., and LEE, J. A.
1966. HETEROSIS IN RELATION TO DEVELOPMENT IN UPLAND COTTON. Crop Sci. 6: 555.GLUSHKOV, N. M., and SKREBTSOV, M. F.
1960. INCREASING THE COTTON CROP THROUGH SATURATION POLLINATION BY BEES.] PchelovodstVo 37(7): 29 - 30. [ln Russian.]GORE, U. R.
1932. DEVELOPMENT OF THE FEMALE GAMETOPHYTE AND EMBRYO IN COTTON. Amer. Jour. Bot. 19: 795 - 807.GROUT, R. A.
1955. HONEY BEES MAKE HYBRID COTTON POSSIBLE. Amer. Bee Jour. 95: 10-11.GULATI, A. N., and TURNER, A. J.
1928. A NOTE ON THE EARLY HISTORY OF COTTON. Indian Cent. Cotton Com. Tech. Bul. 17, 10 pp.GUSEINOV, A. M., and MUKHTAROV, Z. M.
1963. [NEW DATA ON THE EFFECTIVENESS OF INTRAVARIETAL HYBRIDIZATION OF COTTON.] Agrobiologiya 4: 490 - 496. [In Russian. ] Biol. Abs. 45(7): 57028, p. 4681. 1964.HADWICH, F.
1961. [HONEYDEW ON COTTON.] Melliand Textilber. 42(5): 487 - 490. [In German.] AA-603/64.HARLAND, S.C.
1943. BREEDING A COTTON IMMUNE FROM NATURAL CROSSING. Nature 151: 307.______ 1949a. METHODS AND RESULTS OF SELECTION EXPERIMENTS WITH PERUVIAN TANGUIS COTTON. Empire Cotton Growing Rev. 26(3): 163-174.
HARLAND S. C.
1949b. METHODS AND RESULTS OF SELECTION EXPERIMENTS WITH PERUVIAN TANGUIS COTTON Empire Cotton Growing Rev. 26(4): 247-255.HAWKINS, B. S., PEACOCK, H. A., and BALLARD, W. W.
1962. HETEROSIS AND COMBINING ABILITY IN UPLAND COTTON- EFFECT ON YIELD. Crop Sci. 2: 543-546.HUGHES, L. C.
1968. MOTES IN VARIETIES OF GOSSYPIUM BARBADENSE. Empire Cotton Growing Rev. 45(3): 266-280.HUMBERT, E. P., and MOGFORD, J. S.
1927. VARIATION IN CERTAIN LINT CHARACTERS IN A COTTON PLANT AND ITS PROGENY. Tex. Agr. Expt. Sta. Bul. 349, 23 pp.HUTCHINSON, J. B., PANSE, V. G., and GOVANDE, G. K.
1938. STUDIES IN PLANT BREEDING AND TECHNIQUE. IV, THE INHERITANCE OF AGRICULTURAL CHARACTERS IN 3 INTERSTRAIN CROSSES OF COTTON. Indian Jour. Agr. Sci. 8: 757 - 775.IVANOVA-PAROISKAYA, M. I.
1950. [HONEY-BEARING COTTON IN UZBEKISTAN.] Pchelovodstvo 9: 479 - 483. [In Russian.]______ 1956. [NECTAR IN COTTON IN UZBEKISTAN AND ITS POLLINATION BY BEES.] In Krishchunas, I. V., and Gubin, A. F., eds. Pollination of Agricultural Plants. Moskva, Gos. Izd-vo Sel'khoz. Lit-ry, pp. 24-44. [In Russian.]
IYENGAR, N. K.
1938. POLLEN TUBE STUDIES IN GOSSYPIUM. Jour. Genet. 37: 69-106.JANKI, R. P., SINGH, R., PRAKASH, R., and HAQUE, M. F.
1968. FLORAL BIOLOGY OF COTTON (GOSSYPIUM SPP.). Indian Jour. Agr. Sci. 38(3): 528 - 535.JENSEN W. A., and FISHER, D. B.
1968. COTTON EMBRYOGENESIS: THE ENTRANCE AND DISCHARGE OF THE POLLEN TUBE IN THE EMBRYO SAC. Planta 78(2): 158 - 183.JOHANSSON, T. S. K.
1959. TRACKING HONEY BEES IN COTTON FIELDS WITH FLUORESCENT PIGMENTS. Jour. Econ. Ent. 52: 572 - 577.JUSTUS, N., and LEINWEBER, C. L.
1960. A HERITABLE PARTIALLY MALE-STERILE CHARACTER IN COTTON. Jour. Hered. 51: 191-192.MEYER, J. R., and ROUX, J. B.
1963. A PARTIALLY MALE-STERILE CHARACTER IN UPLAND COTTON. Crop Sci. 3: 428-429.KALYANARAMAN, S. M., and SANTHANAM, V.
1957. A REVIEW OF RECENT TRENDS IN COTTON BREEDING TECHNIQUES. Indian Cotton Growing Rev. 11(2): 131 - 135.KAZIEV, T. I.
1955. [ROLE OF BEES IN POLLINATION OF COTTON.] Klopkovodstvo 12: 39 - 40. [ In Russian. ] AA-472/61.______ 1956a. [THE WORKING OF COTTON BY BEES IN WESTERN AZERBAIDZHAN.] Uchen. Zap. Kirovabad. Ped. Inst. (4): 153-156. [In Russian.] AA-210/61.
______ 1956b. [ACTITY OF HONEYBEES ON COTTON.] Sotsial. Sell Khoz. Azerbaidzhan 8: 36-39. [In Russian.] AA-933/63.
KAZIEV, T. I. 1958. [ADDITIONAL POLLINATION OF COTTON BY BEES.] Pchelovodstvo 35(8): 28 - 34. [In Russian.]
______ 1959a. [INFLUENCE OF FERTILIZATION AND WATERING ON NECTAR SECRETION OF COTTON.] Pchelovodstvo 36(1): 25 - 28. [In Russian.]
______ 1959b. [THE INFLUENCE OF CLIMATIC CONDITIONS ON THE PROCESS OF NECTAR SECRETION IN COTTON.] Pchelovodstvo 36(9): 37 - 39. [In Russian.] AA-342/60.
______ 1960. [THE INFLUENCE OF BEE POLLINATION ON THE QUALITY OF COTTON.] PchelovodstVo 37(7): 33 - 35. [In Russian.] AA-333/62.
______ 1961a. [NECTAR SECRETION IN COTTON AND THE ROLE OF BEES IN INCREASING ITS YIELD.] Report of the Inter-University Scientific Conference, pp. 68 - 69: The Universities' Contribution to Agriculture, Moscow. [In Russian.] AA-935/63.
______ 1961b [INFLUENCE OF BEE POLLINATION ON THE PROGENY OF COTTON VARIETIES.] Uchen. Zap. kirovabad ped. Inst. (8): 100 - 112. [In Azerbaidzhani, Russian summary.] AA-934/63.
______ 1963. THE NECTAR YIELD OF THE COTTON PLANT AND THE TASK OF BEES IN INCREASING ITS FERTILITY. In 19th Internatl. Apic. Cong. Proc., Prague, p. 57.
______ 1964. [PROBLEMS OF NECTAR SECRETION IN COTTON AND THE ROLE OF HONEY BEES IN INCREASING ITS YIELD.] 215 pp. Baku, AzerbaidEhan Gos. Izd. [In Russian.]
______ 1967. [SOME AGROTECHNICAL MEASURES EXPEDITING INCREASED NECTAR PRODUCTIVITY OF COTTON.] In 21st Internatl. Apic. Cong. Proc., College Park, Md., Aug. Pp. 452-454. [In Russian.] KEARNEY T. H. 1921. POLLINATION OF PIMA COTTON IN RELATION TO THE YIELD OF SEED AND FIBER. Jour. Hered. 12(3): 99 - 101.
______ 1923. SELF-FERTILIZATION AND CROSS-FERTILIZATION IN PIMA COTTON. U.S. Dept. Agr. Dept. Bul. 1134, 68 pp.
______ 1926. BAGGING COTTON FLOWERS TO PREVENT ACCIDENTAL CROSS- POLLINATION. Jour. Hered. 17: 273 - 279.
______and HARRISON, G. J. 1932. POLLEN ANTAGONISM IN COTTON. Jour. Agr. Res. 44: 191 - 226.
KHAN, H., and AFZAL, M.
1950. VICINISM IN COTTON. Indian Cotton Growing Rev. 4(4): 227-239.KIME, P. H., and TILLEY, R. H.
1947. HYBRID VIGOR IN UPLAND COTTON. Amer. Soc. Agron. Jour. 39: 308 - 317.KNIGHT, R. L., and ROSE, M. F.
1954. A NEW APPROACH TO THE TECHNIQUE OF COTTON SELECTION. Empire Cotton Growing Rev. 31(3): 162-170.KOHEL, R. J.
1968. EFFECT OF GENOTYPE AND HETEROZYGOSITY OF POLLEN SOURCE AND METHOD OF APPLICATION ON SEED SET AND SEED AND FIBER DEVELOPMENT IN COTTON. Crop Sci. 8: 293-295.______and RICHMOND, T. R.
1962. AN EVALUATION OF SEED-YIELD POTENTIAL OF COMPLETELY MALE-STERILE COTTON IN AREAS OF HIGH AND LOW NATURAL CROSSING. Agron. Jour. 54: 525 - 528.______and RICHMOND, T. R.
1969. EVALUATION OF SYNTHETIC VARIETIES OF UPLAND COTTON DEVELOPED UNDER TWO LEVELS OF NATURAL CROSSING. Crop Sci. 9: 647-651.______ 1921. CROSS-FERTILIZATION AND STERILITY IN COTTON. Agr. Jour. India, part I, 16(1): 52 - 59; part II, 16(4): 406 - 409.
______ 1928. CONTINUED SELF-POLLINATION IN COTTON. Nature 122: 314.
KULIEV, A. M.
1958. THE USE OF BEES TO INCREASE COTTON YIELD. In 17th Internatl. Apic. Cong. Proc. Rome. Pp. 65-66.LEE. J. A., MILLER, P. A., and RAWLINGS, J. O.
1967. INTERACTION OF COMBINING ABILITY EFFECTS WITH ENVIRONMENT IN DIALLEL CROSSES OF UPLAND COTTON (G. HIRSUTUM L.). Crop Sci. 7: 477-481.LINSKENS, H. F.
1964. POLLEN PHYSIOLOGY AND FERTILIZATION. 257 pp. Symposium at University of Nijmegen, The Netherlands. Aug. 1963. North Holland Publishing Co., Amsterdam.LLOYD, F. E.
1920. ENVIRONMENTAL CHANGES AND THEIR EFFECT UPON BOLL- SHEDDING IN COTTON. N.Y. Acad. Sci. Ann. 29: 1-131.LODEN, H. D., and RICHMOND, T. R.
1951. HYBRID VIGOR IN COTTON - CYTOGENIC ASPECTS AND PRACTICAL APPLICATIONS. Econ. Bot. 5: 387-408.______LEWIS, C. F., and RICHMOND, T. R.
1950. THE EFFECTS OF TIME AND METHOD OF POLLINATION ON SEED SET IN AMERICAN UPLAND COTTON. Agron. Jour. 42: 560-564.LUKEFAHR, M. J., and MARTIN, D. F.
1964. THE EFFECT OF VARIOUS LARVAL AND ADULT DIETS ON THE FECUNDITY AND LONGEVITY OF THE BOLLWORM, TOBACCO BUDWORM AND COTTON LEAFWORM. Jour. Econ. Ent. 57: 233 - 235.______and RHYNE, C.
1960. EFFECTS OF NECTARILESS COTTON ON POPULATIONS OF THREE LEPIDOPTEROUS INSECTS. Jour. Econ. Ent. 53: 242-244.MAHADEVAN, V., and CHANDY, K. C.
1959. PRELIMINARY STUDIES ON THE INCREASE IN COTTON YIELD DUE TO HONEY BEE POLLINATION. Madras Agr. Jour. 47: 23-26. 188MARANI, A.
1967. HETEROSIS AND COMBINING ABILITY IN INTRASPECIFIC AND INTERSPECIFIC CROSSES OF COTTON. Crop Sci. 7: 519-522.MCGREGOR, S. E.
1958. RELATION OF HONEY BEES TO POLLINATION OF MALE-STERILE COTTON. In Eleventh Cotton Impr. Conf. Proc.: 93-94, Houston.____ 1959. COTTON-FLOWER VISITATION AND PO1LEN DISTRIBUTION BY HONEY BEES. Science 129: 97-98.
____ and TODD, F. E. 1955. COTTON IS BENEFITED BY BEES. Prog. Agr. in Ariz. 7(1): 12.
____ and TODD, F. E. 1956. HONEYBEES AND COTTON PRODUCTION. Gleanings Bee Cult. 84: 649 - 652, 701.
____ RHYNE, C., WORLEY, S., JR., and TODD, F. E.
1955. THE ROLE OF HONEY BEES IN COTTON POLLINATION. Agron. Jour. 47: 23-25.MCMILLIAN, W. W.
1959. THE POTENTIAL POLLINATORS OF COTTON INMONOKOVA FLORENCE AND DARLINGTON COUNTIES,1955. SOUTH CAROLINA. 56 pp. M.S. Thesis, Clemson Agr. Col.MCNAMARA, H. C., HOOTON, D. R., and PORTER. D. D.
1940. DIFFERENTIAL GROWTH RATES IN COTTON VARIETIES AND THEIR RESPONSE TO SEASONAL CONDITIONS AT GREENVILLE, TEXAS U.S. Dept. Agr. Tech. Bul. 710, 44 ppMEADE, R. M.
1918. BEEKEEPING MAY INCREASE THE COTTON CROP. Jour. Hered. 9: 282 - 285.MELL P. H.
1894. EXPERIMENTS IN CROSSING FOR THE PURPOSE OF IMPROVING THE COTTON FIBER. Ala. Agr. Expt. Sta. Bul. 56, 47 pp.MEL'NICHENKO, A. N.
1963. [BEES THEMSELVES INCREASE THE NECTAR PRODUCTIVITY OF FLOWERS.] Pchelovodstvo 40(9): 32-35. tIn Russian.] Biol. Abs. 45(7): 62074, p. 5069, 1964.MERCADO MESA, O.
1956. [AUTOGAMY AND THE EFFECT OF INSECTICIDES ON THE FERTILITY OF TANGUIS COTTON.] Lima Estac. Expt. Agr. de La Molina. lnforme Mens. 30 (346): 30- 36. [In Spanish.]MEYER, J. R., and MEYER, V. G.
1961. ORIGIN AND INHERITANCE OF NECTARILESS COTTON. Crop Sci. 1: 167-169.____ Roux, J. B., and THOMAS, R. O.
1958. HYBRID COTTON IS AIM OF RESEARCH PROJECT IN DELTA. Miss. Farm Res. 21(5): 2.MEYER, V. G.
1969. SOME EFFECTS OF GENES, CYTOPLASM, AND ENVIRONMENT ON MALE STERILITY OF COTTON (GOSSYPIUM). Crop Sci. 9: 237-242.____ and MEYER, J. R.
1965. CYTOPLASMICALLY CONTROLLED MALE STERILITY IN COTTON. Crop Sci. 5: 444.MILLER, P. A., and LEE, J. A.
1964. HETEROSIS AND COMBINING ABILITY IN VARIETAL TOP CROSSES OF UPLAND COTTON, G. HIRSUTUM L. Crop Sci. 4: 646-649.MINKOV, S. G.
1953a. [HONEYBEES AND COTTON.] Pchelovodstvo (8): 41-44. [In Russian. ] AA-210/54.____ 1953b. [ON VARIETAL ENTOMOPHILOUS CHARACTERISTICS IN COTTON.] KhlopkovodstVo 3(7): 40-44. [In Russian.]
____ 1956. [NECTAR PRODUCTIVITY OF COTTON AND THE ROLE OF BEES IN CROSS-POLLINATION.] Trud. Kazakh. Op. Stants. Pchelovodstva 1: 109-150. [In Russian.] AA-363/68.
____ 1957. NECTAR PRODUCTIVITY OF COTTON IN SOUTH-KAZAKH REGION. Pchelovodstvo 34(12): 35-40. AA-190/59.
MIRAVALLE, R. J.
1964. A NEW BULKED-POLLEN METHOD FOR COTTON CROSS- POLLINATION. Jour. Hered. 55: 276-280.MONOKOVA, F. N., and CHEBOTNIKOVA, K. M.
1955. [AN INCREASE IN THE NECTAR PRODUCTIVITY OF HONEY PLANTS BY MEANS OF VARIOUS FERTILIZERS.] Pchelovodstvo 8: 44 - 46. [In Russian.] Biol. Abs. 31: 2119, July, 1957.MOUND, L. A.
1962. EXTRA-FLORAL NECTARIES OF COTTON AND THEIR SECRETIONS. Empire Cotton Growing Rev. 39: 254 - 261.MURAMOTO, H.
1958. THE GENETIC COMBINING ABILITY OF CERTAIN VARIETIES OF GOSSYPIUM HIRSUTUM AS MEASUR] FOR AGRONOMIC AND SPINNING QUALITIES. 119 pp. Dissertation submitted to University of Arizona as partial fulfillment of requirements for Ph.D. degree, University of Arizona Library, Tucson.O'KELLY, J. F.
1942. DEGENERATION WITHIN COTTON VARIETIES Agron. Jour. 34: 782 - 796.PARKS, H. B.
1921. THE COTTON PLANT AS A SOURCE OF NECTAR. Amer. Bee Jour. 61: 391.PATEL, G. B., and PATEL, C. I.
1952. LONG STAPLE COTTON FROM HYBRID SEED OF CROSS BETWEEN HIRSUTUM AND BARBADENSE SPECIES. Indian Cotton Growing Rev. 6(4): 205-210.PEARSON, N. L.
1949a. MOTE TYPES IN COTTON AND THEIR OCCURRENC AS RELATED TO VARIETY, ENVIRONMENT, POSITION IN LOCK, LOCK SIZE AND NUMBER OF LOCKS PER BOLL. U.S. Dept. Agr. Tech. Bul. 1000, 37 pp.____ 1949b. FALSE MOTES IN COTTON, THEIR ORIGIN, DESCRIPTION AND VARIATION IN NUMBER. Jour. Agr. Res. 78: 705 - 717.
PEEBLES, R. H.
1942. PURE SEED PRODUCTION OF EGYPTIAN-TYPE COTTON. U.S. Dept. Agr. Cir. 646, 20 pp.____ 1956. FIRST ATTEMPT TO PRODUCE HYBRID COTTONSEED. Amer. Bee Jour. 96: 51 - 52, 75.
PORTER, D. D.
1936. POSITIONS OF SEEDS AND MOTES IN LOCKS AND LENGTHS OF COTTON FIBERS FROM BOLLS BORNE AT DIFFERENT POSITIONS ON PLANTS AT GREENVILLE, TEXAS. U.S. Dept. Agr. Tech. Bul. 509, 13 pp.RADOEV, L.
1963. STUDIES ON BEE POLLINATION AND HONEY PRODUCTIVITY OF COTTON. In 19th Internatl. Apic. Cong. Proc., Liblice, Czechoslovakia. P. 99.____ 1965. [BEE POLLINATION OF COTTON.] Pchelovodstvo 9: 39 - 41. [In Russian.]
____ and BOZHINOV, M. 1961. [A STUDY ON THE NECTAR SECRETION OF COTTON AND THE ROLE OF BEES IN ITS POLLINATION.] Izv. Kompl. sel. lnst. Chirpan 1: 87 - 108. [In Bulgarian, English summary.]
REA, H. E.
1928. LOCATION OF MOTES IN THE UPLAND COTTON LOCK. Amer. Soc. Agron. Jour. 20: 1064-1068.____ 1929. VARIETAL AND SEASONAL VARIATION OF MOTES IN UPLAND COTTON. Amer. Soc. Agron. Jour. 21: 481 - 486, 1154-1155.
REED, E. L.
1917. LEAF NECTARIES OF GOSSYPIUM. Bot. Gaz. 63: 229-231.RICHMOND, T. R., and LEWIS, C. F.
1951. EVALUATION OF VARIETAL MIXTURES OF COTTON. Agron. Jour. 43: 66-70.RICKS, J. R., and BROWN, H. B.
1916. COTTON EXPERIMENTS. Miss. Agr. Expt. Sta. Bul. 178, 40 pp.ROHM AND HAAS CO.
1958. FW-450 A SELECTIVE GAMETOCIDE. Information sheet, January. Washington Square, Philadelphia.ROSE, M. F., and HUGHES, L. C.
1955. PROGRESS REPORT OF THE COTTON BREEDING STATION, SEASON OF 1954-55. Cesira Research Station, Wad Medani, pp. 2-12.SAAKYAN, T. A.
1962. [AN EMBRYOLOGICAL STUDY OF THE PROCESS OF FERTILIZATION OF COTTON WITH DIFFERENT METHODS OF POLLINATION.] Biol. Nauki (AN Arm. SSR), 15(7): 59 - 65. [In Russian.] Biol. Abs. 45(6): 52655, p. 4273. 1964.SAPPENFIELD, W. P.
1963. ESTIMATES OF NATURAL CROSSING IN UPLAND COTTON IN SOUTHEAST MISSOURI. Crop Sci. 3: 566.SEN, K. R. D., and AFZAL, M.
1937. VARIATION IN THE CHARACTERS OF COTTON IN RELATION TO THE POSITION OF BOLLS ON THE PLANT. Indian Jour. Agr. Sci. 7(1): 35 - 47.SHISHIKIN, E. A.
1946. [HONEY BEES IN THE SERVICE OF COTTON POLLINATION.] PchelovodstVo 23(5-6): 31-32. [In Russian.] Biol. Abs. 21(2): 26151, p. 2551, 1947.SHISHIKIN, E. A.
1952. [EFFECT OF POLLINATION BY HONEYBEES ON INCREASING THE PRODUCTIVITY OF COTTON.] In Krishchunas, I. V., and Gubin, A. F., eds. [Pollination of agricultural plants.]. Moskva, Gos. Izd-vo Selkhoz Lit-ry, pp. 95 - 103. [In Russian.]SHOEMAKER, D. N.
1911. NOTES ON VICINISM IN COTTON IN 1908. Amer. Breeders' Assoc. 6: 252 - 254.SIDHU, A. S., and SINGH, S.
1961. STUDIES ON AGENTS OF CROSS POLLINATION IN COTTON. Indian Cotton Growing Rev. 15: 341-353.____ 1962. ROLE OF HONEYBEES IN COTTON PRODUCTION. Indian Cotton Growing Rev. 16: 18 - 23.
SIMPSON, D. M.
1948. HYBRID VIGOR FROM NATURAL CROSSING FOR IMPROVING COTTON PRODUCTION. Amer. Soc. Agron. Jour. 40: 970 - 979.____ 1954a. NATURAL CROSS-POLLINATION IN COTTON. U.S. Dept. Agr. Tech Bul, 1094, 17 pp.
____ 1954b. "RADICAL METHODS" SEEK HYBRID COTTON VIGOR. Tenn. Farm and Home Sci. 9: 4.
____ and DUNCAN, E. N.
1953. EFFECT OF SELECTING WITHIN SELFED LINES ON THE YIELD AND OTHER CHARACTERS OF COTTON. Agron. Jour. 45: 275-279.____ and DUNCAN, E. N.
1956. COTTON POLLEN DISPERSAL BY INSECTS. Agron. Jour. 48: 305 - 308.SKREBTSOV, M F.
1964. [THE PROBLEM OF ABUNDANT POLLINATION OF COTTON BY HONEY BEES.l Trud. nauch.-issled Inst. Pchelov. 246-264. [In Russian, English summary.] AA-864/65.STEPHENS, S. G.
1956. THE COMPOSITION OF AN OPEN POLLINATED SEGREGATING COTTON POPULATION. Amer. Nat. 90(850): 25 - 39.____ and FINKNER, M. D.
1953. NATURAL CROSSING IN COTTON. Econ. Bot. 7(3): 257 - 269.STITH, L. S.
1910. A BEE IN HYBRID COTTON PRODUCTION? In The Indispensable Pollinators, Ark. Agr. Ext. Serv. Misc. Pub. 127, pp. 28 - 32.STROMAN, G. N.
1961. AN APPROACH TO HYBRID COTTON AS SHOWN BY INTRA AND INTERSPECIFIC CROSSES. Crop Sci. 1: 363-366.TER-AVANESYAN, D. V.
1952. [BIOLOGY OF POLLINATION AND FLOWERING IN COTTON.] Bul. Appl. Bot., Genet. and Plant Breed. 29(2): 149-169. [In Russian.] Plant Breed. Abs. 24: 3155. 1954.____ 1959. THE EFFECT OF QUANTITY OF POLLEN USED ON THE INHERITANCE OF CHARACTERS. Indian Jour. Genet. and Plant Breed. 19(1): 30-35
____ and LALAEV, G. B. 1954. [INTRAVARIETAL CROSSING OF COTTON PLANTS SOWN AT DIFFERENT TIMES.] Agrobiologiya 6: 57-61.1 In Russian.] Biol. Abs. 31(2): 15023, p. 1481, 1957. 190
THAKAR, B. J., and SHETH, D. S.
1955. COMMERCIAL PRODUCTION OF HYBRID SEED IN BOMBAY STATE. Indian Cotton Growing Rev. 9(3): 120-122.THEIS, S. A.
1953. AGENTS CONCERNED WITH NATURAL CROSSING OF COTTON IN OKLAHOMA. Amer. Soc. Agron. Jour. 45: 481 - 484.THOMSON, N. J.
1966. COTTON VARIETY TRIALS IN THE ORD VALLEY, NORTHWESTERN AUSTRALIA. Empire Cotton Growing Rev. 43(1): 18-21.TRELEASE, W.
1879. NECTAR, WHAT IT IS AND SOME OF ITS USES. In Comstock, J. H., U.S. Dept. Agr. Rpt. on Cotton Insects, pt. 3, pp. 319-343.TRUSHKIN, A. V.
1956. [WAYS OF IMPROVING THE HEREDITARY CHARACTERISTICS OF COTTON SEED WITH A NEW SYSTEM OF KOLKHOZ SEED RAISING.] Nauchn. Tr. Uzb. Sel'khoz. Inst. 9(1): 75-85. [In Russian.] Biol. Abs. 35(3): 27822, p. 2454, 1960.____ 1960a. [THE USE OF BEES FOR THE POLLINATION OF COTTON.] Agrobiologiya 5: 787-788. [In Russian.] AA-335l62.
____ 1960b. [BEE POLLINATION INCREASES YIELD OF COTTON.] KhlopkovodstVo 10(12): 33-34. [In Russian.]
____ and TRUSKINA, L. P. 1964. [REPEATED POLLINATION BY BEES INCREASES VIGOR OF COTTON.] Khlopkovodsho 6: 30-31. [In Russian.] Plant Breed. Abs. 36: 2313, p. 308, 1966.
TSYGANOV, S. K.
1953. [REMARKS ON THE POLLINATING ACTIVITY OF HONEY BEES.] Uzbekistan Akad. Nauk Inst. Zool. i. Parasitol.i927 Trudy 1: 91-122. [In Russian.]TURNER, J. H.
1948. HYBRID VIGOR STUDIES. Ga. Coastal Plain Expt. Sta. Ann. Rpt. Bul. 46, pp. 18-20.____ 1953a. A STUDY OF HETEROSIS IN UPLAND COTTON. 1. YIELD OF HYBRIDS COMPARED WITH VARIETIES. II. COMBINING ABILITY AND INBREEDING EFFECTS. Agron. Jour. 45: 484 - 486, 487 - 490.
____ 1953b. DIFFERENTIAL RESPONSE OF COTTON VARIETIES TO NATURAL CROSSINGS. Agron. Jour. 45: 246-248.
____ 1959. HYBRID COTTON BREEDING PROGRAM. Calif. Agr. 13(3): 3, 15.
TURNER, J. H., and MIRAVALLE, R. J.
1961. 'HYBRID COTTON' FROM ACALA-4-42 COULD BOOST YIELD AND QUALITY BY 10% ON 60'S. Cotton Digest 33 (31): 18-19.TYLER, F. J.
1908. THE NECTARIES OF COTTON. In Brown, E., Scofield, C. S., Hedgcock, G. G., an others, U.S. Dept. Agr. Bul. 131, pt. 5, pp. 45 - 56.UNITED STATES DEPARTMENT OF AGRICULTURE.
1971. COTTON VARIETIES PLANTED 1967-1971. U.S. Dept. Agr. Consumer and Market. Serv.. Cotton Div., Memphis, 32 pp.VANSELL, G. H.
1944a. COTTON NECTAR IN RELATION TO BEE ACTIVITY AND HONEY PRODUCTION. Jour. Econ. Ent. 37: 528-530.____ 1944b. SOME WESTERN NECTARS AND THEIR CORRESPONDING HONEYS. Jour. Econ. Ent. 37: 530-532.
WAFA, A. K., and IBRAHIM, S. H.
1957. THE HONEYBEE AS AN IMPORTANT INSECT FOR POLLINATION. Cairo University, Faculty Agr., Bul. 162, 13 pp.____ and IBRAHIM, S. H.
1959. POLLINATORS OF THE CHIEF SOURCES OF NECTAR AND POLLEN GRAIN PLANTS IN EGYPT. Soc. Ent. Egypt Bul. 43: 133 - 154.____ and IBRAHIM, S. H.
1960. THE EFFECT OF THE HONEYBEE AS A POLLINATING AGENT ON THE YIELD OF CLOVER AND COTTON. Cairo University, Faculty Agr., Bul. 206, 44 pp.WANJURA, D. F., HUDSPETH, E. B., JR., and BILBRO, J. D., JE
1969. EMERGENCE TIME, SEED QUALITY, AND PLANTIN DEPTH EFFECTS ON YIELD AND SURVIVAL OF COTTON (G. HIRSUTUM L.). Agron. Jour. 61: 63-65.WARE, J. O.
1927. COTTON SPACING. Ark. Agr. Expt. Sta. Bul. 220, 80 pp.____ 1931. COTTON BREEDING, SPACING, AND VARIETY STUDIES. Ark. Agr. Expt. Sta. 43d Ann. Rpt. for fiscal year ending June 30, 1931, pp. 30 - 31.
WEAVER, J. B., JR., and ASHLEY, T.
1971. ANALYSIS OF A DOMINANT GENE FOR MALE STERILITY IN UPLAND COTTON. Crop Sci. 11: 596 - 597.WEBBER, H. J.
1903. IMPROVEMENT OF COTTON BY SEED SELECTION. U.S. Dept. Agr. Yearbook 1902, pp. 365-386.WYKES, G. R.
1952. THE PREFERENCE OF HONEY BEES FOR SOLUTIONS OF VARIOUS SUGARS. Jour. Expt. Biol. 29: 511-518.